Introduction
Chronological age accounts for only a fraction of the substantial inter‑individual variability observed in health span, physical function, and susceptibility to chronic disease, prompting growing interest in biomarkers that more accurately reflect biological aging. Among the many proposed candidates, glucose homeostasis is particularly compelling because disturbances in glycemic control emerge early, are quantifiable with high temporal resolution, and are mechanistically linked to multiple aging pathways, including oxidative stress, advanced glycation, endothelial dysfunction, and low‑grade inflammation.
Glucose metabolism deteriorates progressively with age, even in individuals without overt diabetes, with population data showing decade‑on‑decade increases in 2‑hour oral glucose tolerance test (OGTT) values and a higher prevalence of postprandial hyperglycemia, while fasting plasma glucose may remain within conventional reference ranges. These post‑challenge and postprandial abnormalities are associated with subclinical vascular damage, increased arterial stiffness, microvascular complications, and higher risk of cardiovascular events, as well as structural and functional brain changes that include gray matter atrophy and cognitive decline. Such findings position glycemic control as a central determinant of vascular, neurological, and multi‑organ aging, beyond its traditional framing as a diabetes‑centric risk factor.
Emerging evidence further suggests that dynamic glycemic metrics, particularly glycemic variability and the magnitude and frequency of postprandial excursions may capture aspects of iological aging more sensitively than static indices such as fasting glucose or glycated hemoglobin (HbA1c). Continuous glucose monitoring (CGM) studies in older yet non‑diabetic adults demonstrate higher mean glucose and hyperglycemia‑related indices compared with younger adults, despite similar fasting levels, while multi‑scale variability measures have been linked to oxidative stress, gray matter loss, and long‑term functional outcomes. Together, these observations support the concept that maintaining a “youthful” glycemic profile, characterized by lower mean glucose, limited postprandial excursions, and reduced variability may serve as a clinically relevant biomarker of biological youth and a target for preventive and longevity-oriented interventions.
Why “Youth” is Metabolic
Youthful physiology is characterized by high insulin sensitivity, low visceral adiposity, and tightly regulated postprandial glucose excursions, which together support efficient substrate utilization and minimal metabolic stress even in the context of intermittent dietary excess. In younger adults, relatively greater skeletal muscle mass, preserved mitochondrial function, and more favourable adipose tissue distribution facilitate rapid clearance of postprandial glucose and limit exposure to hyperglycemia‑induced oxidative stress and glycation. These features contribute to a metabolic milieu that protects vascular, neural, and other organ systems, aligning “youth” with a state of metabolic flexibility and resilience [1,2,3,4].
With advancing age, shifts in body composition, including increased visceral and ectopic fat, reduced lean mass, and adipose tissue dysfunction converge with declines in physical activity and alterations in beta‑cell responsiveness to impair glucose regulation. Older adults commonly exhibit higher mean glucose levels and exaggerated postprandial excursions, even when fasting glucose remains within conventionally normal limits, reflecting an early dysglycemic state driven by insulin resistance and impaired first‑phase insulin secretion. Epidemiologic data demonstrate that isolated postprandial hyperglycemia in the presence of normal fasting glucose and HbA1c confers a substantially increased risk of cardiovascular morbidity and mortality, underscoring the prognostic importance of post‑challenge glycemia in aging populations [1,2,5,6,7].
These age‑related metabolic changes underpin a broad spectrum of downstream consequences, including higher rates of cardiovascular disease, neurodegeneration, and functional decline. Insulin resistance and postprandial hyperglycemia are linked to endothelial dysfunction, arterial stiffness, and microvascular damage, while also associating with impaired cerebral glucose uptake, increased amyloid burden, and greater risk of cognitive impairment and Alzheimer’s disease. Collectively, these observations position metabolic health and specifically the preservation of insulin sensitivity, low visceral adiposity, and stable postprandial glucose, as a core anchor of both perceived and objectively measured youthfulness across the lifespan [1,3,8,9,10,11].
From Fasting Glucose to Dynamic Glycemic Metrics
Traditional markers of glucose homeostasis, such as fasting plasma glucose and glycated hemoglobin (HbA1c), capture only limited dimensions of glycemic exposure and often underestimate early dysglycemia in aging populations. Individuals can display normal fasting glucose and HbA1c while already exhibiting impaired glucose tolerance and pronounced postprandial hyperglycemia on oral glucose tolerance testing (OGTT), a pattern that carries increased risk for cardiovascular events and mortality. These discrepancies highlight that static, fasting‑based measures fail to fully reflect the dynamic nature of daily glucose fluctuations and may miss subtle, age‑related impairments in glycemic control that are clinically relevant [1,5,12].
Continuous glucose monitoring (CGM) provides high‑resolution characterization of 24‑hour glucose profiles and enables derivation of composite metrics such as time in range (TIR), mean glucose, coefficient of variation (CV), and various risk and control indices including glucose management indicator (GMI), glycemic risk assessment in diabetes equation (GRADE), M‑value, and J‑index. Studies in adults without diabetes show that these CGM‑based parameters can distinguish “younger” from “older” glycemic phenotypes, even when standard laboratory tests remain within reference limits, revealing age‑related changes in the distribution of glucose values across the day. For example, older but otherwise healthy adults demonstrate higher mean and minimum glucose, elevated hyperglycemia‑related indices (e.g., high blood glucose index and GRADE hyperglycemia), and higher composite control scores (GMI, M‑value, J‑index) compared with younger adults, despite broadly similar time in range and variability metrics [13,14].
Continuous glucose monitoring (CGM) provides high‑resolution characterization of 24‑hour glucose profiles and enables derivation of composite metrics such as time in range (TIR), mean glucose, coefficient of variation (CV), and various risk and control indices including glucose management indicator (GMI), glycemic risk assessment in diabetes equation (GRADE), M‑value, and J‑index. Studies in adults without diabetes show that these CGM‑based parameters can distinguish “younger” from “older” glycemic phenotypes, even when standard laboratory tests remain within reference limits, revealing age‑related changes in the distribution of glucose values across the day. For example, older but otherwise healthy adults demonstrate higher mean and minimum glucose, elevated hyperglycemia‑related indices (e.g., high blood glucose index and GRADE hyperglycemia), and higher composite control scores (GMI, M‑value, J‑index) compared with younger adults, despite broadly similar time in range and variability metrics [13,14].
Glycemic Variability and Vascular Aging
Short‑ and intermediate‑term glycemic variability have been shown to correlate closely with oxidative stress and the accumulation of advanced glycation end products (AGEs), mechanistically linking glucose fluctuations to endothelial dysfunction and accelerated vascular aging. In a recent cross‑sectional study of patients with type 2 diabetes undergoing continuous glucose monitoring, coefficient of variation (representing short‑term variability) and mean of daily differences (representing intermediate‑term variability) were independent predictors of oxidative stress as measured by diacron‑reactive oxygen metabolites, as well as AGE accumulation assessed by skin autofluorescence, which in turn associated with the presence of vascular complications. Experimental data support this association, with landmark studies demonstrating that oscillating glucose produces greater impairment in endothelial function and higher levels of nitrotyrosine, a marker of oxidative stress, compared with high constant high glucose exposure over 24 hours, even when total glucose exposure is equivalent or lower. These effects appear to be mediated by repeated cycles of hyperglycemia‑induced oxidative burst that do not fully resolve during normoglycemic intervals, creating a cumulative vascular injury that exceeds the damage from stable hyperglycemia alone [15,16,17,18,19].

Elevated glycemic variability is robustly associated with higher cardiovascular risk in populations with diabetes, and multiple pathophysiologic pathways, including oxidative stress, inflammation, AGE formation, and glycation are plausible mediators of this risk at earlier stages of dysglycemia. Postprandial hyperglycemia, characterized by acute glucose excursions, induces rapid generation of reactive oxygen species, upregulation of inflammatory adhesion molecules such as ICAM‑1, and activation of leukocyte integrins, which together promote endothelial dysfunction, arterial stiffness, and atherogenesis. Epidemiologic evidence further indicates that postprandial glucose spikes are stronger predictors of cardiovascular mortality than fasting glucose or HbA1c, with elevated 2‑hour post‑load glucose conferring increased risk even among individuals classified as having normal glucose tolerance. The observation that glycemic variability, quantified by mean amplitude of glycemic excursion (MAGE), correlates more strongly with urinary markers of oxidative stress than does mean glucose itself underscores the independent atherogenic contribution of glucose fluctuations beyond average glycemic exposure [5,8,16,18,20,21].
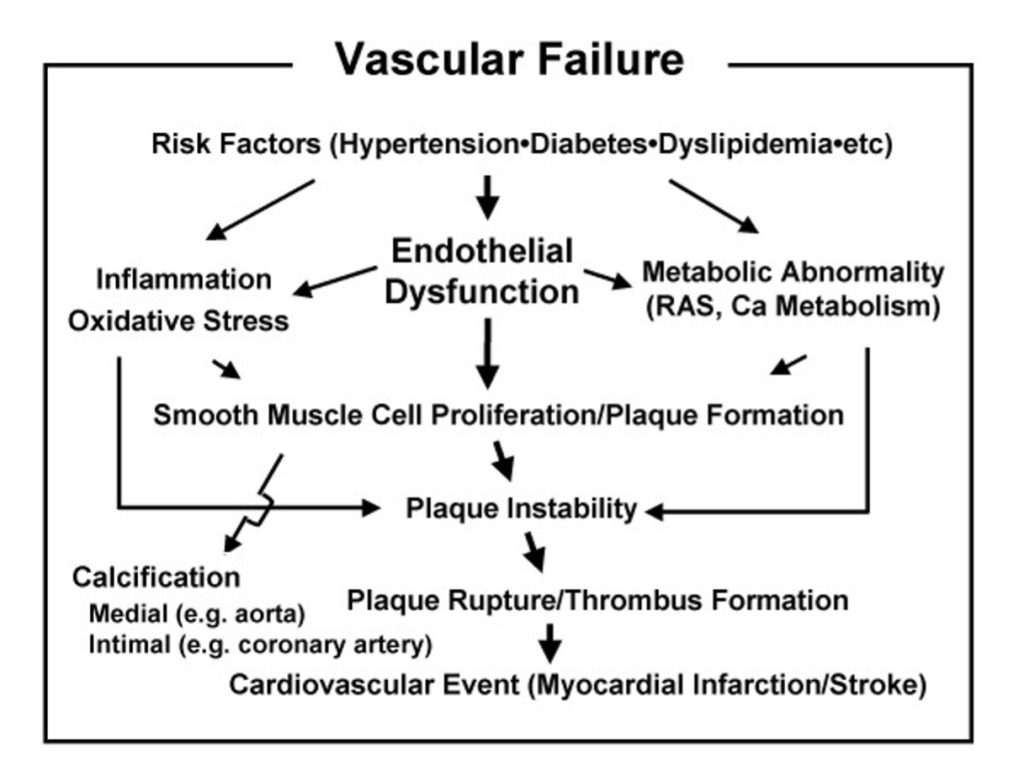
Minimizing large postprandial excursions and day‑to‑day glycemic variability may therefore represent an anti‑aging strategy at the vascular level that complements and potentially exceeds the benefit of simply lowering mean glucose. Interventions that reduce postprandial spikes, whether through dietary modification, incretin‑based therapies, or physical activity have been shown to improve flow‑mediated vasodilation and reduce oxidative stress markers, supporting the concept that targeting glucose dynamics can attenuate vascular aging processes. Furthermore, the prolonged metabolic memory effect of glycemic excursions, whereby inflammatory gene expression persists for days after a single hyperglycemic spike, suggests that sustained optimization of glucose stability could yield cumulative vascular protection over years and decades, making glycemic variability a priority target in preventive and longevity medicine [6,8,16,17,18,21].
Glucose Control and Brain Structure/ Function
In adolescents with type 1 diabetes, tighter blood sugar control has been shown to improve gray and white matter characteristics and cognitive performance, partially normalizing brain maturation trajectories that are otherwise disrupted by chronic dysglycemia. A randomized controlled trial evaluating automated insulin delivery (closed-loop) technology against standard care over six months found that teenagers using closed-loop systems achieved significantly better glycemic control, with more time in target glucose range, and demonstrated superior performance on perceptual reasoning tasks compared with conventionally managed controls. Brain imaging revealed that teens with improved glucose control exhibited lower cortical gray matter volume, reduced subcortical tissue volume, and increased white matter integrity, changes consistent with more efficient neural pruning and myelination that characterize healthy adolescent neurodevelopment. Importantly, these structural and functional improvements occurred within only six months, and were associated with degree of glycemic control independent of treatment modality, suggesting that normalizing glucose dynamics can attenuate or even reverse some diabetes-related brain abnormalities during critical developmental windows [22,23,24,25].
In older adults with type 2 diabetes, multi-scale glycemic variability has been independently associated with gray matter atrophy in limbic and temporo-parietal regions and with poorer cognitive performance, implicating glucose fluctuations as a distinct driver of brain aging beyond mean glucose or HbA1c. A landmark study employing continuous glucose monitoring and advanced signal processing techniques demonstrated that individuals with greater short-term glycemic variability (cycles of 0.5–2.0 hours) exhibited reduced gray matter volumes in key structures including the cingulum, insula, hippocampus, and temporal lobes, as well as worse performance on cognitive tests, independent of HbA1c and hypoglycemic episode frequency. These relationships were time-scale-dependent, with higher-frequency glucose oscillations most strongly linked to structural changes in the central autonomic network and cognitive domains such as learning and memory, suggesting that the pattern and magnitude of glucose swings exert specific pathophysiologic effects on brain structure and function that differ from those of sustained hyperglycemia [26].
Long-term glycemic variability, as measured by visit-to-visit coefficient of variation in fasting plasma glucose and HbA1c, has been linked to increased risk of cognitive decline, dementia, and Alzheimer’s disease in middle-aged and elderly individuals with diabetes, suggesting that stabilizing glucose levels over years and decades may help preserve cognitive “youth.” Prospective cohort studies show that higher variability in HbA1c and fasting glucose is associated with greater incidence of all-cause dementia and Alzheimer’s disease, with effects that persist after adjustment for mean glucose levels, hypoglycemia, and other cardiovascular risk factors. In one large cohort, middle-aged and elderly patients with higher long-term glucose variability demonstrated significantly worse cognitive function across multiple domains, with HbA1c standard deviation emerging as the strongest predictor of mild cognitive impairment. These findings underscore that both the average level of glucose exposure and the amplitude of fluctuations contribute independently to neurodegenerative risk, and collectively support the concept that maintaining stable, near-normal glucose dynamics throughout adulthood is a key determinant of cognitive resilience and brain health in later life [27,28,29,30,31].
Glycemic Trajectories Across the Lifespan
Population data from large cross-sectional and longitudinal studies demonstrate that 2-hour post-load glucose during oral glucose tolerance testing (OGTT) rises steadily with each decade of adult life, peaking around the seventh decade, while fasting plasma glucose can remain deceptively stable or increase only modestly over the same period, highlighting the primacy of postprandial control in the pathogenesis of age-related glucose intolerance. Among individuals who never develop overt type 2 diabetes, fasting glucose increases progressively but gradually across the lifespan in both nonhuman primates and humans, whereas the 2-hour OGTT value shows more pronounced age-related deterioration, reflecting impaired beta-cell responsiveness and declining insulin sensitivity that manifest most clearly in the post-challenge state. This dissociation between fasting and postprandial measures underscores a fundamental limitation of relying on fasting glucose or HbA1c alone for early detection of age-related metabolic decline, as individuals can maintain normal fasting values while already exhibiting significant impairment in glucose tolerance and elevated cardiovascular risk [1,5,32,33,34].
Even in healthy older adults without diabetes, continuous glucose monitoring (CGM) reveals higher mean glucose and hyperglycemia-related indices compared with young adults, despite similar time in range and variability measures, indicating a subtle but pervasive upward shift in the entire glucose distribution with aging. In a recent controlled study comparing healthy young (mean age ~24 years) and older (mean age ~70 years) adults over five consecutive days of CGM, older participants exhibited significantly higher mean glucose (6.25 vs 5.89 mmol/L), higher minimum glucose (3.70 vs 3.19 mmol/L), and elevated hyperglycemia-related scores (HBGI, GRADE hyperglycemia), while variability metrics (coefficient of variation, MAGE, MODD) and time in range remained statistically similar between groups. Importantly, glycemic control indices, including glucose management indicator (GMI), M-value, and J-index were all significantly higher in older adults, reflecting poorer overall control despite broadly similar glucose fluctuations, which suggests that the absolute level of glucose exposure, rather than its variability, differentiates younger from older metabolic phenotypes in the absence of disease [14].
These findings collectively support the concept that a “younger” metabolic phenotype is characterized by lower mean glucose, lower minimum glucose, and smaller postprandial excursions throughout adult life, with preservation of these characteristics potentially serving as both a marker and a mediator of healthy aging. Longitudinal CGM data from adults without diabetes document a gradual age-related shift from glucose values predominantly in the 70–99 mg/dL range toward the 100–139 mg/dL range, paralleling the decade-by-decade rise in 2-hour OGTT glucose observed in population studies. The observation that older adults maintain good glycemic control and low variability yet still exhibit upward-shifted glucose profiles suggests that aging-related changes in insulin sensitivity, beta-cell function, and tissue glucose uptake create a new metabolic “set point” that, while not overtly pathologic, may nonetheless contribute to accelerated vascular, cognitive, and functional decline over time. Consequently, interventions aimed at preserving a youthful glucose trajectory through diet, physical activity, pharmacotherapy, or other means may represent a foundational strategy for extending health span and delaying biological aging [1,4,13,14,35].
Positioning CGM Metrics as Youth Biomarkers
Continuous glucose monitoring (CGM) enables high-resolution, real-world assessment of glucose dynamics across the full spectrum of daily living, translating discrete behaviours including diet, sleep, physical activity, and stress into quantifiable, time-stamped metrics that reflect an individual’s metabolic resilience and regulatory capacity. Unlike single fasting or post-challenge measurements, CGM captures the entirety of glycemic excursions and fluctuations over days to weeks, providing insight into both the magnitude and frequency of postprandial spikes, nocturnal glucose stability, and the temporal relationships between lifestyle exposures and glucose responses. Studies in healthy young adults under standardized conditions demonstrate that CGM can discriminate the effects of specific foods, aerobic versus anaerobic exercise, and psychosocial stress on glucose patterns, revealing substantial inter-individual variability even among metabolically healthy individuals. Importantly, this capacity to track glucose responses in response to everyday contexts allows CGM to function not merely as a diagnostic tool but as a continuous biofeedback system that surfaces actionable information about metabolic health, supporting behaviour modification and personalized optimization strategies well before any clinical pathology emerges [36,37,38,39,40].
In adults with type 2 diabetes, machine learning models trained on continuous glucose monitoring data alone or in combination with manually logged meals and medication intake can predict next-in-time postprandial glucose (PPG) excursions with substantial accuracy, reinforcing the clinical utility and feasibility of dynamic, personalized glycemic metrics. A recent study of individuals with type 2 diabetes found that personalized models achieved a mean F1-score of approximately 76% for predicting PPG excursions using only six days of individual CGM data, with some individuals reaching 100% predictive accuracy, illustrating that glucose responses are sufficiently patterned and repeatable to enable anticipatory dietary and pharmacologic interventions. Incorporating additional data streams such as manual meal logs, physical activity, and temporal variables, modestly improves prediction performance for some individuals, while for others CGM data alone is sufficient, highlighting the heterogeneity of postprandial vulnerability and the necessity of individualized modelling approaches. These findings demonstrate that predicting and attenuating glucose spikes through personalized nutrition and behaviour recommendations is not only feasible but can be automated and refined iteratively, opening the door to “just-in-time” interventions that prevent hyperglycemic excursions before they occur and thereby limit cumulative glycemic stress over the lifespan [40,41,42,43].
Incorporating CGM-derived metrics such as time in range (TIR), mean glucose, coefficient of variation, and composite control indices into multi-dimensional “biological age” panels could meaningfully strengthen risk stratification and intervention targeting in preventive and longevity medicine. Biological aging clocks based on routine clinical biomarkers have been shown to predict disease risk, mortality, and subjective health status with greater accuracy than chronological age alone, and the addition of dynamic glucose measures would enrich these models by capturing a domain, metabolic flexibility and postprandial control that is both modifiable and mechanistically linked to vascular, neurologic, and functional aging. For instance, individuals whose CGM profiles reveal higher mean glucose, elevated glycemic variability, or frequent excursions beyond target ranges may exhibit accelerated biological aging even in the absence of overt diabetes, warranting earlier or more intensive lifestyle, nutritional, or pharmacologic interventions. Moreover, the longitudinal monitoring capabilities of CGM allow for repeated, low-burden assessment of glycemic trajectories over time, enabling clinicians and researchers to track responses to interventions, detect early deviations from “youthful” glucose patterns, and adjust therapeutic strategies dynamically. By reframing glucose dynamics as a core component of biological youth and integrating CGM metrics into composite aging biomarker platforms, the field can move toward more personalized, data-driven approaches to health span extension and age-related disease prevention [14,38,44,45,46].

Mechanistic Links: From Glucose to Aging Hallmarks
Recurrent hyperglycemia promotes the formation of advanced glycation end products (AGEs), oxidative stress, mitochondrial dysfunction, and low-grade systemic inflammation, the core of hallmarks of aging that collectively accelerate vascular and tissue damage across multiple organ systems. Chronic exposure to elevated glucose drives non-enzymatic glycation of proteins, lipids, and nucleic acids, generating AGEs that bind to receptors such as RAGE (receptor for advanced glycation end products), triggering sustained NF-κB activation and perpetuating inflammatory cascades. Simultaneously, hyperglycemia increases glucose flux through the mitochondrial electron transport chain, leading to excessive reactive oxygen species (ROS) production that overwhelms endogenous antioxidant defenses, impairs mitochondrial redox balance, and induces further mitochondrial dysfunction in a self-amplifying cycle. Hyperglycemia also activates multiple deleterious metabolic pathways, including the polyol pathway, protein kinase C (PKC) signalling, and hexosamine biosynthetic pathway, each of which independently contributes to oxidative stress, depletion of critical antioxidants like glutathione, and activation of pro-inflammatory mediators. These interconnected processes culminate in widespread cellular injury, accelerated senescence, and tissue dysfunction that are characteristic of biological aging, with evidence demonstrating that mitochondrial dysfunction serves as a central hub linking oxidative stress, inflammation, and the aging phenotype [8,19,47,48,49,50,51].
Postprandial glucose spikes exert particularly potent adverse effects on vascular and neural tissues, impairing endothelial function, increasing arterial stiffness, and exacerbating neuroinflammation, thereby linking glycemic instability directly to cardiovascular and brain aging phenotypes. Acute postprandial hyperglycemia rapidly induces oxidative stress and upregulates inflammatory adhesion molecules such as ICAM-1, activates leukocytes, and triggers sustained NF-κB-driven gene expression in the vasculature that persists for days even after glucose returns to normal, a phenomenon termed “metaboli memory.” Experimental models demonstrate that oscillating glucose produces greater endothelial dysfunction and higher oxidative stress markers (e.g., nitrotyrosine) than constant hyperglycemia at equivalent total glucose exposure, underscoring the uniquely deleterious nature of glucose fluctuations. In clinical studies, postprandial hyperglycemia increases brachial pulse wave velocity, a marker of arterial stiffness in in patients with type 2 diabetes, and epidemiologic data consistently show that post-challenge glucose levels predict cardiovascular mortality more strongly than fasting glucose or HbA1c. In the brain, chronic hyperglycemia disrupts the blood-brain barrier, facilitates amyloid-beta accumulation, and activates the NLRP3 inflammasome, triggering caspase-1-dependent release of synaptotoxic IL-1β and pyroptosis, mechanistically linking metabolic dysregulation to neurodegeneration and accelerated cognitive aging [5,8,17,18,51,52,53,54].
Insulin resistance and beta-cell stress arising from chronic glycemic load further propagate metabolic inflexibility, sarcopenia, and adipose tissue dysfunction, creating a self-reinforcing aging loop that integrates multiple organ systems and physiologic domains. Insulin resistance impairs skeletal muscle glucose uptake and protein anabolism, directly contributing to muscle fiber atrophy, reduced muscle strength, and sarcopenia, while simultaneously reducing insulin-responsive tissue mass and thereby worsening systemic insulin resistance in a vicious cycle. Obesity and visceral adiposity, which both drive and result from insulin resistance, promote secretion of pro-inflammatory cytokines (TNF-α, IL-6) and adipokines that further impair insulin signalling at the level of insulin receptor substrate (IRS) proteins, activate stress kinases (JNK, IKKβ), and exacerbate oxidative stress and mitochondrial dysfunction in muscle, liver, and adipose tissue. Chronic hyperglycemia and lipid overload induce beta-cell glucotoxicity and lipotoxicity, leading to increased ROS production, ER stress, activation of apoptotic pathways, and ultimately beta-cell failure and impaired insulin secretion, which accelerates the transition from compensated insulin resistance to overt diabetes. Importantly, aging itself is associated with increased visceral fat, mitochondrial dysfunction, oxidative stress, and chronic low-grade inflammation, all of which predispose to insulin resistance while insulin resistance accelerates sarcopenia, adipose dysfunction, and beta-cell senescence, creating an integrated metabolic aging phenotype in which glycemic dysregulation is both cause and consequence of broader physiologic decline [55,56,57].

Lifestyle and Nutritional Levers to “Re-Youth” Glucose
Meal-based dietary interventions, particularly those emphasizing low-energy and lower-glycemic-load patterns, significantly improve glucose regulation in adults with obesity and type 2 diabetes and likely attenuate postprandial spikes in high-risk non-diabetic individuals through multiple complementary mechanisms. Low-glycemic-index (low-GI) and low-glycemic-load (low-GL) diets reduce the rate of carbohydrate digestion and absorption, leading to flatter postprandial glucose curves, decreased insulin demand, and improved beta-cell preservation. A meta-analysis and systematic review of randomized controlled trials consistently demonstrate that low-GI/GL diets lower HbA1c and postprandial glucose in individuals with type 2 diabetes, independent of significant weight loss, suggesting direct metabolic benefits beyond caloric restriction. Importantly, even within the low-GI range, the absolute GI value matters: consumption of an ultralow-GI biscuit (GI 24) compared with a moderate-low-GI biscuit (GI 54) resulted in 56% lower postprandial glucose and 45% lower insulin response at breakfast in healthy adults monitored by continuous glucose monitoring, with benefits persisting at a subsequent meal, illustrating the physiologic relevance of fine-tuning glycemic load. Mechanistically, low-GL meals enhance satiety, reduce ghrelin suppression, lower inflammatory markers, and may improve whole-body insulin sensitivity, though data on the latter remain mixed and warrant further large-scale trials. Practically, adopting low-GI/GL dietary patterns involves selecting intact whole grains, legumes, non-starchy vegetables, and foods enriched with soluble fiber, plant-based fats, and protein, while minimizing refined carbohydrates and high-GI starches, a strategy that is feasible, scalable, and directly translates into measurable reductions in glycemic excursions and variability in real-world settings [58,59,60,61].
Resistance and aerobic training improve insulin sensitivity, reduce visceral adiposity, and flatten glucose excursions, making structured exercise a cornerstone “youth-preserving” intervention for glycemic control with distinct but synergistic physiologic effects. Resistance training augments insulin sensitivity by approximately 48% as measured by the gold-standard euglycemic-hyperinsulinemic clamp, primarily by increasing skeletal muscle mass and quality, which is responsible for roughly 80% of insulin-mediated glucose uptake, thereby directly addressing the muscle insulin resistance that characterizes aging and type 2 diabetes. High-resistance exercise programs are particularly effective at reducing visceral adipose tissue, a key driver of systemic insulin resistance and inflammation and also lower total cholesterol, LDL cholesterol, triglycerides, and blood pressure in older adults with type 2 diabetes. Aerobic exercise complements these effects by lowering fasting and postprandial glucose, reducing HbA1c (meta-analysis: -0.66% independent of weight loss), and decreasing average glucose concentrations by approximately 14.4 mg/dL as assessed by continuous glucose monitoring. Concurrent (combined aerobic plus resistance) training produces the greatest metabolic benefits, with reductions in HbA1c of -0.34% to -0.59% compared with either modality alone, alongside improvements in lipid profiles and blood pressure, underscoring the additive and possibly synergistic nature of these exercise modes. Practically, an effective “re-youthing” exercise prescription might include 3 sessions per week of progressive resistance training (2–3 sets of 8–12 repetitions at 60–80% one-repetition maximum targeting major muscle groups) combined with 150 minutes per week of moderate-to-vigorous aerobic activity (e.g., brisk walking, cycling, swimming at 60–75% maximal heart rate), with emerging evidence suggesting that afternoon or fed-state exercise may minimize acute hyperglycemic responses compared with fasted morning sessions, particularly for resistance modalities [62,63,64].
Sleep optimization, stress management, and circadian alignment further modulate glucose dynamics via neuroendocrine pathways, though high-quality interventional data remain comparatively limited and mechanistic understanding is still evolving. Acute sleep deprivation (24 hours to 5 days) decreases insulin sensitivity, impairs fasting and postprandial glucose tolerance, and reduces pancreatic beta-cell insulin secretion, effects mediated by elevated evening cortisol (promoting gluconeogenesis and hepatic glucose output), increased sympathetic tone and catecholamine release (inducing insulin resistance), and systemic low-grade inflammation. Circadian misalignment such as occurs with shift work or mistimed eating independently mpairs glucose tolerance and beta-cell function, leading to elevated postprandial glucose excursions even when sleep duration is preserved, with mechanisms involving altered melatonin and cortisol rhythms relative to the sleep-wake and feeding-fasting cycles. Specifically, when meals occur during the biological night, glucose and insulin levels are significantly higher due to reduced insulin sensitivity and impaired pancreatic responsiveness driven by circadian phase, with genome-wide association studies linking circadian clock genes, glucose homeostasis, and pancreatic melatonin signalling. Practically, interventions to “re-youth” glucose through sleep and circadian optimization include maintaining consistent sleep-wake schedules (ideally 7–9 hours per night), avoiding late-night eating (aligning feeding to daylight hours), minimizing exposure to artificial light at night (to preserve melatonin rhythms), and managing psychosocial stress through mindfulness, yoga, or cognitive-behavioural strategies that lower cortisol and sympathetic activation. While these lifestyle levers are biologically plausible and supported by mechanistic and short-term experimental data, long-term randomized controlled trials are needed to definitively establish their efficacy for preserving youthful glucose profiles and preventing age-related metabolic decline in diverse populations [65,66,67,68].

Clinical and Digital Health Applications
Framing glycemic control as a biomarker of youth enables earlier identification of at‑risk individuals who present with normal fasting glucose yet display exaggerated postprandial excursions or elevated glycemic variability on continuous glucose monitoring (CGM). Large cohort studies show that isolated post‑challenge hyperglycemia, even in the absence of fasting hyperglycemia, predicts cardiovascular events and mortality, indicating that conventional screening can miss individuals with clinically relevant dysglycemia whose fasting indices remain in range. CGM exposes these “hidden” phenotypes by quantifying time above range, magnitude and frequency of postprandial spikes, and day‑to‑day variability, allowing clinicians to classify patients whose dynamic glucose profiles resemble an “older” vascular and metabolic age despite apparently normal standard tests. This repositions glycemic monitoring from a purely diagnostic tool for established diabetes to an early warning system for accelerated cardiometabolic and biological aging in ostensibly healthy or preclinical populations [1,5,13,14,69,70,71,72,73].
AI‑driven analytics can leverage CGM, diet, activity, and wearable data to derive individualized glycemic aging scores, predict postprandial responses, and recommend tailored micro‑interventions in near real time. Personalized machine learning models trained on approximately six days of individual CGM (with or without meal and medication logs) achieve F1‑scores around 73–76% for predicting forthcoming postprandial glucose excursions in adults with type 2 diabetes, with some individuals reaching 100% predictability, demonstrating that PPG patterns are sufficiently structured to support anticipatory guidance. Integration of smartwatch‑derived signals (e.g., sleep, heart rate variability, activity) further improves PPG prediction accuracy to the 84–86% range in healthy individuals, highlighting the added value of multimodal data streams for capturing behavioral and physiologic drivers of glycemic instability. These predictive models can be embedded into digital therapeutics and just‑in‑time adaptive interventions (JITAIs) that deliver context‑aware prompts, such as adjusting meal composition, timing a short walk, or modifying medication timing when an individual enters a personalized “state of vulnerability” for glucose spikes, thereby reducing cumulative glycemic stress without necessitating continuous clinician oversight [41,43,74,75].
Integrating CGM‑derived metrics into preventive and longevity clinics offers a path toward more precise, behaviorally anchored strategies to maintain youthful metabolic profiles over decades. Contemporary biological age clocks built from routine clinical biomarkers, metabolomics, or DNA methylation patterns already outperform chronological age for predicting morbidity, mortality, and functional decline, and emerging models explicitly incorporate cardiometabolic and inflammatory markers to derive interpretable “metabolic age” scores. Adding CGM metrics such as time in range, mean glucose, indices of hyperglycemia, and measures of short‑ and long‑term glycemic variability would enrich these platforms by capturing dynamic aspects of metabolic flexibility and postprandial control that link directly to vascular, neurocognitive, and functional aging. Clinically, individuals could be stratified by glycemic age relative to their chronological age (e.g., “glucose age +8 years”), with targeted interventions (nutrition, exercise, pharmacotherapy, sleep and circadian optimization) titrated until CGM‑based metrics shift toward a more “youthful” profile, providing both a therapeutic target and an objective feedback loop for long‑term health span management [13,14,44,45,69,76,77,78].
Limitations and Research Gaps
Most mechanistic links between glycemic variability and aging have been elucidated in populations with established diabetes, limiting the direct extrapolation of these findings to non‑diabetic individuals across the full adult age range. Long‑term prospective cohorts that combine continuous glucose monitoring with detailed phenotyping in metabolically “healthy” adults remain relatively scarce, creating uncertainty about the precise exposure, response relationships between subtle glycemic instability and downstream aging phenotypes in the general population [13,14,15,26,45,69].
At present, there is no universally accepted “glycemic age” construct, nor standardized thresholds that define youthful versus aged glucose dynamics in otherwise healthy adults. Existing biological age clocks rarely incorporate dynamic glycemic metrics, and proposed CGM‑based cut‑offs for time in range, variability, or postprandial excursion magnitude are largely derived from diabetes care frameworks rather than from aging‑focused normative data [13,14,44,77,79].
Future research should integrate CGM‑derived metrics with multi‑omics (including metabolomics, proteomics, and epigenetic clocks), advanced imaging (vascular, cardiac, and neuroimaging), and functional outcomes (cognition, physical performance, frailty indices) to rigorously test glycemic control as a core biomarker within composite biological age models. Such studies will be essential to establish causal pathways, define age‑ and sex‑specific reference ranges for “youthful” glycemic patterns, and determine whether targeted interventions that normalize glucose dynamics can meaningfully decelerate biological aging and extend health span [44,45,77,78].
Conclusion
Across youth, midlife, and older age, tighter blood sugar control operationalized as lower mean glucose levels, restrained postprandial excursions, and reduced glycemic variability is consistently associated with more favourable vascular, brain, and cognitive profiles. Data from both diabetic and non‑diabetic populations indicate that individuals who maintain more stable glycemic patterns exhibit less endothelial dysfunction, reduced markers of oxidative stress, and slower progression of structural brain changes, aligning tighter glycemic control with preserved organ integrity and delayed functional decline. Within this framework, glycemic dynamics can be viewed not only as risk factors for cardiometabolic disease but also as integral readouts of systemic physiological “youth.”
Conceptualizing dynamic glycemic metrics as a biomarker of youth reframes glucose monitoring from a reactive tool used mainly in advanced diabetes to a proactive signal for detecting early deviations in metabolic and biological aging trajectories. Continuous glucose monitoring and related analytic approaches enable high‑resolution characterization of individual glucose responses to diet, physical activity, sleep, and stress, allowing clinicians and researchers to identify subclinical dysglycemia long before traditional thresholds are met. This shift supports the integration of glycemic patterns into composite biological age models and risk scores that can guide earlier, more personalized intervention strategies.
Embedding this glycemic‑youth paradigm into AI‑enabled health technologies and clinical practice may open a new frontier in longevity medicine, in which preserving a “youthful” glycemic signature becomes a central therapeutic aim rather than a secondary outcome. Machine‑learning models trained on CGM, lifestyle, and multi‑omics data can help predict postprandial responses, estimate glycemic aging trajectories, and generate individualized behavioural or nutritional prescriptions targeted at flattening glucose excursions and reducing variability. As these tools mature and become more accessible, they have the potential to transform glucose metrics into actionable, everyday biomarkers of youth, bridging the gap between mechanistic aging science and real‑world preventive care.
Reference
- Chia CW, Egan JM, Ferrucci L. Age-Related Changes in Glucose Metabolism, Hyperglycemia, and Cardiovascular Risk. Circulation Research. 2018 Sep 14;123(7):886–904.
- Mancuso P, Bouchard B. The Impact of Aging on Adipose Function and Adipokine Synthesis. Frontiers in Endocrinology [Internet]. 2019 Mar 11;10. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6421296/
- Lalia AZ, Dasari S, Johnson ML, Robinson MM, Konopka AR, Distelmaier K, et al. Predictors of Whole-Body Insulin Sensitivity Across Ages and Adiposity in Adult Humans. The Journal of Clinical Endocrinology & Metabolism. 2016 Feb 1;101(2):626–34.
- Palmer AK, Jensen MD. Metabolic changes in aging humans: current evidence and therapeutic strategies. Journal of Clinical Investigation [Internet]. 2022;132(16). Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9374375/
- Gerich JE. Clinical Significance, Pathogenesis, and Management of Postprandial Hyperglycemia. Archives of Internal Medicine. 2003 Jun 9;163(11):1306.
- Shibib L, Mo Al-Qaisi, Guess N, Miras AD, Greenwald SE, Pelling M, et al. Manipulation of Post-Prandial Hyperglycaemia in Type 2 Diabetes: An Update for Practitioners. Diabetes Metabolic Syndrome and Obesity. 2024 Aug 1;Volume 17:3111–30.
- Chandler-Laney PC, Phadke R, Granger WM, Fernández JR, Muñoz AJ, Man CD, et al. Age-related changes in insulin sensitivity and β-cell function among European American and African American women. Obesity (Silver Spring, Md) [Internet]. 2011 Mar 1;19(3):528. Available from: https://www.ncbi.nlm.nih.gov/labs/pmc/articles/PMC3074467/
- Hanssen NMJ, Kraakman MJ, Flynn MC, Nagareddy PR, Schalkwijk CG, Murphy AJ. Postprandial Glucose Spikes, an Important Contributor to Cardiovascular Disease in Diabetes? Frontiers in Cardiovascular Medicine. 2020 Sep 18;7.
- Barone E, Di Domenico F, Perluigi M, Butterfield DA. The interplay among oxidative stress, brain insulin resistance and AMPK dysfunction contribute to neurodegeneration in type 2 diabetes and Alzheimer disease. Free Radical Biology and Medicine. 2021 Nov;176:16–33.
- de la Monte SM. Insulin Resistance and Neurodegeneration: Progress Towards the Development of New Therapeutics for Alzheimer’s Disease. Drugs. 2016 Dec 17;77(1):47–65.
- Willette AA, Bendlin BB, Starks EJ, Birdsill AC, Johnson SC, Christian BT, et al. Association of Insulin Resistance With Cerebral Glucose Uptake in Late Middle–Aged Adults at Risk for Alzheimer Disease. JAMA Neurology [Internet]. 2015 Sep 1;72(9):1013. Available from: https://jamanetwork.com/journals/jamaneurology/fullarticle/2398420
- Mongraw-Chaffin M, Bertoni AG, Golden SH, Mathioudakis N, Sears DD, Szklo M, et al. Association of Low Fasting Glucose and HbA1c With Cardiovascular Disease and Mortality: The MESA Study. Journal of the Endocrine Society. 2019 Mar 1;3(5):892–901.
- Couselo M, Portos-Regueiro C, González-Rodríguez M, Manuel García-Lopez J, Alonso-Sampredro M, Rodríguez-González R, et al. Aging of glucose profiles in an adult population without diabetes. Diabetes Research and Clinical Practice. 2022 Jun;188:109929.
- Köhlmoos A, Dittmar M. Glycemic variability and control by CGM in healthy older and young adults and their relationship with diet. Journal of the Endocrine Society [Internet]. 2025 May 8 [cited 2025 Oct 10];9(7). Available from: https://doi.org/10.1210/jendso/bvaf081
- Ohara M, Takahashi N, Nobuaki Takehana, Osaka N, Sugita H, Michishige Terasaki, et al. Association of glycemic variability with oxidative stress and AGE accumulation in type 2 diabetes. Scientific Reports [Internet]. 2025 Dec 11 [cited 2026 Jan 12]; Available from: https://www.nature.com/articles/s41598-025-31845-x
- Offler LJ, Wells LK, Palmer TM. The Impact of Glycaemic Variability on Vascular Dysfunction in Diabetes. Biomolecules [Internet]. 2025 Nov 3;15(11):1544–4. Available from: https://www.mdpi.com/2218-273X/15/11/1544?utm_source=chatgpt.com
- Ceriello A, Esposito K, Piconi L, Ihnat MA, Thorpe JE, Testa R, et al. Oscillating Glucose Is More Deleterious to Endothelial Function and Oxidative Stress Than Mean Glucose in Normal and Type 2 Diabetic Patients. Diabetes. 2008 Feb 25;57(5):1349–54.
- Node K, Inoue T. Postprandial hyperglycemia as an etiological factor in vascular failure. Cardiovascular Diabetology [Internet]. 2009;8(1):23. Available from: https://cardiab.biomedcentral.com/articles/10.1186/1475-2840-8-23
- Nowotny K, Jung T, Höhn A, Weber D, Grune T. Advanced Glycation End Products and Oxidative Stress in Type 2 Diabetes Mellitus. Biomolecules [Internet]. 2015 Mar 16;5(1):194–222. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4384119/
- Koska J, Saremi A, Howell S, Bahn G, De Courten B, Ginsberg H, et al. Advanced Glycation End Products, Oxidation Products, and Incident Cardiovascular Events in Patients With Type 2 Diabetes. Diabetes Care. 2017 Dec 5;41(3):570–6.
- Jarvis PRE, Cardin JL, Nisevich-Bede PM, McCarter JP. Continuous glucose monitoring in a healthy population: understanding the post-prandial glycemic response in individuals without diabetes mellitus. Metabolism. 2023 Sep 1;146(155640):155640.
- Mauras N, Buckingham B, White NH, Tsalikian E, Weinzimer SA, Jo B, et al. Impact of Type 1 Diabetes in the Developing Brain in Children: A Longitudinal Study. Diabetes Care [Internet]. 2021 Feb 10;44(4):983–92. Available from: https://diabetesjournals.org/care/article/44/4/983/138591/Impact-of-Type-1-Diabetes-in-the-Developing-Brain
- digitale @stanford.edu img src=’https://med stanford edu/news/media-contacts/erin_digitale/_jcr_content/image img 620 high jpg/digitale-erin-90 jpg’ alt=’Erin DEDED is the pediatrics science writer in the O of CE her at. Improved blood sugar control helps normalize diabetic teens’ brains, Stanford-led study finds [Internet]. News Center. Available from: https://med.stanford.edu/news/all-news/2022/08/teen-diabetes-brain.html
- Stanisławska-Kubiak M, Katarzyna Anna Majewska, Agata Krasińska, Wais P, Majewski D, Mojs E, et al. Brain functional and structural changes in diabetic children. How can intellectual development be optimized in type 1 diabetes? Therapeutic advances in chronic disease. 2024 Jan 1;15.
- In Type 1 Diabetes, Glycemic Control in Adolescence Can Stave Off Neurological Damage [Internet]. HCP Live. 2022. Available from: https://www.hcplive.com/view/type-1-diabetes-glycemic-control-in-adolescence-can-stave-off-neurological-damage\\
- Cui X, Abduljalil A, Manor BD, Peng CK, Novak V. Multi-Scale Glycemic Variability: A Link to Gray Matter Atrophy and Cognitive Decline in Type 2 Diabetes. Gong G, editor. PLoS ONE. 2014 Jan 24;9(1):e86284.
- Underwood PC, Zhang L, Mohr DC, Prentice JC, Nelson RE, Budson AE, et al. Glycated Hemoglobin ATime in Range and Dementia in Older Adults With Diabetes. JAMA Network Open [Internet]. 2024 Aug 2;7(8):e2425354–4. Available from: https://jamanetwork.com/journals/jamanetworkopen/fullarticle/2821878 1c
- Ballesteros S, Redondo MT. Is glycemic variability associated with a risk of Alzheimer’s disease in older adults with diabetes mellitus? Journal of Laboratory and Precision Medicine. 2017;2:90–0.
- Ding J, Shi Q, Tao Q, Su H, Du Y, Pan T, et al. Correlation between long-term glycemic variability and cognitive function in middle-aged and elderly patients with type 2 diabetes mellitus: a retrospective study. PeerJ. 2023 Dec 20;11:e16698–8.
- Yu J, Lee KN, Kim HS, Han K, Lee SH. Cumulative effect of impaired fasting glucose on the risk of dementia in middle-aged and elderly people: a nationwide cohort study. Scientific Reports [Internet]. 2023 Nov 23;13(1):20600. Available from: https://www.nature.com/articles/s41598-023-47566-y#Sec2\\
- Ding J, Shi Q, Tao Q, Su H, Du Y, Pan T, et al. Correlation between long-term glycemic variability and cognitive function in middle-aged and elderly patients with type 2 diabetes mellitus: a retrospective study. PeerJ. 2023 Dec 20;11:e16698–8.
- Jagannathan R, Neves JS, Dorcely B, Chung ST, Tamura K, Rhee M, et al. The Oral Glucose Tolerance Test: 100 Years Later. Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy [Internet]. 2020 Oct;Volume 13:3787–805. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7585270/
- Palliyaguru DL, Shiroma EJ, Nam JK, Duregon E, Vieira Ligo Teixeira C, Price NL, et al. Fasting blood glucose as a predictor of mortality: Lost in translation. Cell Metabolism. 2021 Nov;33(11):2189-2200.e3.
- Shapiro AJ, Holden EC, McGovern PG, Alderson D, Morelli SS. Comparison of 2-Hour Oral Glucose Tolerance Test and Hemoglobin A1C in the Identification of Pre-Diabetes in Women with Infertility and Recurrent Pregnancy Loss. Clinical Medicine Insights: Reproductive Health. 2019 Jan;13:117955811983128.
- Palliyaguru DL, Shiroma EJ, Nam JK, Duregon E, Vieira Ligo Teixeira C, Price NL, et al. Fasting blood glucose as a predictor of mortality: Lost in translation. Cell Metabolism. 2021 Nov;33(11):2189-2200.e3.
- Kinny F, Läer S, Obarcanin E, Kinny F, Läer S, Obarcanin E. Continuous Glucose Monitoring under standardised conditions regarding diet, exercise and stress in Healthy Young People (CGM-HYPE study): An exploratory clinical trial. PLOS Digital Health [Internet]. 2025 Nov 14;4(11):e0001087–7. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC12617953/
- Attia P. Continuous glucose monitoring to improve health in non-diabetics [Internet]. Peter Attia. 2023. Available from: https://peterattiamd.com/cgm-in-non-diabetics/
- CGM-Led Multimodal Tracking with Chatbot Support: An Autoethnography in Sub-Health [Internet]. Arxiv.org. 2025 [cited 2026 Jan 12]. Available from: https://arxiv.org/html/2510.25381v1
- Should I Use Continuous Glucose Monitoring (CGM)? | PatientsEngage [Internet]. PatientsEngage. 2025. Available from: https://www.patientsengage.com/conditions/should-i-use-continuous-glucose-monitoring-cgm
- Gutierrez-Osuna R, Kerr D, Mortazavi B, Das A. CGMacros: a scientific dataset for personalized nutrition and diet monitoring [Internet]. Physionet.org. 2025. Available from: https://physionet.org/content/cgmacros/1.0.0/
- Brügger V, Kowatsch T, Jovanova M. Predicting postprandial glucose excursions to personalize dietary interventions for type-2 diabetes management. PubMed [Internet]. 2025 Jul 17;15(1):25920–0. Available from: https://www.nature.com/articles/s41598-025-08003-4
- Xiong X, Xue Y, Cai Y, He J, Su H. Prediction of personalised postprandial glycaemic response in type 1 diabetes mellitus. Frontiers in Endocrinology. 2024 Jul 9;15.
- Choudhry NK, Priyadarshini S, Swamy J, Mehta M. Use of Machine Learning to Predict Individual Postprandial Glycemic Responses to Food Among Individuals With Type 2 Diabetes in India: Protocol for a Prospective Cohort Study. JMIR Research Protocols [Internet]. 2025 Jan 23;14:e59308. Available from: https://www.researchprotocols.org/2025/1/e59308
- Meyer DH, Mejia G, Molière A, Panupong Wangprapa, Tanawat Khunlertkit, Polakit Teekakirikul, et al. A sex-adjusted 7-biomarker clinical aging clock for translational preventative medicine. Scientific Reports [Internet]. 2025 Dec 10 [cited 2026 Jan 12];15(1):43538–8. Available from: https://www.nature.com/articles/s41598-025-27478-9
- Mahdi Moqri, Herzog C, Poganik JR, Justice JN, Belsky DW, Higgins-Chen A, et al. Biomarkers of aging for the identification and evaluation of longevity interventions. Cell. 2023 Aug 1;186(18):3758–75.
- Top 5 longevity biomarkers: tests to extend your healthspan • #1 Continuous Glucose Monitoring App Australia | Vively [Internet]. Vively.co.nz. 2025 [cited 2026 Jan 12]. Available from: https://www.vively.co.nz/post/top-5-longevity-biomarkers-tests-to-extend-your-healthspan
- González P, Lozano P, Ros G, Solano F. Hyperglycemia and Oxidative Stress: An Integral, Updated and Critical Overview of Their Metabolic Interconnections. Hyperglycemia and Oxidative Stress: An Integral, Updated and Critical Overview of Their Metabolic Interconnections [Internet]. 2023 May 27;24(11):9352–2. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC10253853/
- Sibony RW, Segev O, Dor S, Raz I. Overview of oxidative stress and inflammation in diabetes. Journal of Diabetes. 2024 Oct 1;16(10).
- Xu X, Pang Y, Fan X. Mitochondria in oxidative stress, inflammation and aging: from mechanisms to therapeutic advances. Signal Transduction and Targeted Therapy [Internet]. 2025 Jun 11;10(1). Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC12159213/
- Giri B, Dey S, Das T, Sarkar M, Banerjee J, Dash SK. Chronic hyperglycemia mediated physiological alteration and metabolic distortion leads to organ dysfunction, infection, cancer progression and other pathophysiological consequences: An update on glucose toxicity. Biomedicine & Pharmacotherapy [Internet]. 2018;107(1):306–28. Available from: https://www.sciencedirect.com/science/article/pii/S0753332218322406
- Raza A, Saleem S, Imran S, Rahman S, Haroon M, Razzaq A, et al. From metabolic dysregulation to neurodegenerative pathology: the role of hyperglycemia, oxidative stress, and blood-brain barrier breakdown in T2D-driven Alzheimer’s disease. Metabolic Brain Disease. 2025 Sep 26;40(7).
- Gordin D, Markku Saraheimo, Jaana Tuomikangas, Aino Soro-Paavonen, Forsblom C, Paavonen K, et al. Influence of Postprandial Hyperglycemic Conditions on Arterial Stiffness in Patients With Type 2 Diabetes. The Journal of Clinical Endocrinology & Metabolism [Internet]. 2016 Jan 5 [cited 2026 Jan 12];101(3):1134–43. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC4803174/
- Aspenani M, Rezaee Z. Oxidative Stress and Metabolic Memory in Diabetes: Focus on the Role of Exercise. Zahedan Journal of Research in Medical Sciences [Internet]. 2026 Jan 4 [cited 2026 Jan 12];28(2). Available from: https://brieflands.com/journals/zjrms/articles/169323
- Ogiso K, Shayo SC, Kawade S, Hashiguchi H, Deguchi T, Nishio Y. Repeated glucose spikes and insulin resistance synergistically deteriorate endothelial function and bardoxolone methyl ameliorates endothelial dysfunction. Bader M, editor. PLOS ONE. 2022 Jan 24;17(1):e0263080.
- Wang M, Tan Y, Shi Y, Wang X, Liao Z, Wei P. Diabetes and Sarcopenic Obesity: Pathogenesis, Diagnosis, and Treatments. Frontiers in Endocrinology. 2020 Aug 25;11.
- Flack KD, Davy KP, Hulver MW, Winett RA, Frisard MI, Davy BM. Aging, Resistance Training, and Diabetes Prevention. Journal of Aging Research. 2011;2011:1–12.
- Zhao X, An X, Yang C, Sun W, Ji H, Lian F. The crucial role and mechanism of insulin resistance in metabolic disease. Frontiers in Endocrinology [Internet]. 2023 Mar 28;14:1149239. Available from: https://pubmed.ncbi.nlm.nih.gov/37056675/#:~:text=Insulin resistance (IR) plays a
- Brownley KA, Heymen S, Hinderliter AL, Galanko J, MacIntosh B. Low-Glycemic Load Decreases Postprandial Insulin and Glucose and Increases Postprandial Ghrelin in White but Not Black Women. The Journal of Nutrition. 2012 May 30;142(7):1240–5.
- Gerontiti E, Shalit A, Katerina Stefanaki, Paraskevi Kazakou, Karagiannakis DS, Melpomeni Peppa, et al. The role of low glycemic index and load diets in medical nutrition therapy for type 2 diabetes: an update. Hormones. 2024 May 16;23.
- Kaur B, Koh M, Ponnalagu S, Henry CJ. Postprandial blood glucose response: does the glycaemic index (GI) value matter even in the low GI range? Nutrition & Diabetes [Internet]. 2020 May 1;10(1). Available from: https://www.nature.com/articles/s41387-020-0118-5
- Jeong Y, Lee E, Park YJ, Kim Y, Kwon O, Kim Y. A Review of Recent Evidence from Meal-Based Diet Interventions and Clinical Biomarkers for Improvement of Glucose Regulation. Preventive Nutrition and Food Science [Internet]. 2020 Mar 31;25(1):9–24. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7143015/
- Syeda A, Battillo D, Visaria A, Malin SK. The Importance of Exercise for Glycemic Control in Type 2 Diabetes. American Journal of Medicine Open [Internet]. 2023;9(9):100031. Available from: https://www.sciencedirect.com/science/article/pii/S2667036423000018
- Lewis C, Rafi E, Dobbs B, Barton T, Hatipoglu B, Malin SK. Tailoring Exercise Prescription for Effective Diabetes Glucose Management. The Journal of clinical endocrinology and metabolism [Internet]. 2025 Spring;dgae908. Available from: https://pubmed.ncbi.nlm.nih.gov/39836084/
- Liu D, Zhang Y, Wu Q, Han R, Cheng D, Wu L, et al. Exercise‐induced improvement of glycemic fluctuation and its relationship with fat and muscle distribution in type 2 diabetes. Journal of diabetes. 2024 Apr 1;16(4).
- Depner CM, Stothard ER, Wright KP. Metabolic Consequences of Sleep and Circadian Disorders. Current Diabetes Reports [Internet]. 2014 May 10;14(7). Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4308960/
- Morris CJ, Yang JN, Garcia JI, Myers S, Bozzi I, Wang W, et al. Endogenous circadian system and circadian misalignment impact glucose tolerance via separate mechanisms in humans. Proceedings of the National Academy of Sciences of the United States of America [Internet]. 2015 Apr 28;112(17):E2225–34. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4418873/
- Mesarwi O, Polak J, Jun J, Polotsky VY. Sleep Disorders and the Development of Insulin Resistance and Obesity. Endocrinology and Metabolism Clinics of North America [Internet]. 2013 Sep;42(3):617–34. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3767932/
- Lin X. The Effects of Sleep on Glycemic Control: Understanding the Connection. J Dia Med Care 2024 [Internet]. 2024 [cited 2025 Jun 10];7(5):269–71. Available from: https://www.openaccessjournals.com/articles/the-effects-of-sleep-on-glycemic-control-understanding-the-connection.pdf
- Belli M, Bellia A, Sergi D, Barone L, Lauro D, Barillà F. Glucose variability: a new risk factor for cardiovascular disease. Acta Diabetologica. 2023 Jun 21;60(10):1291–9.
- Liarakos AL, Panagiotou G, Chondronikola M, Wilmot EG. Continuous Glucose Monitoring in People at High Risk of Diabetes and Dysglycaemia: Transforming Early Risk Detection and Personalised Care. Life [Internet]. 2025 Oct 10 [cited 2025 Nov 15];15(10):1579–9. Available from: https://www.mdpi.com/2075-1729/15/10/1579
- Joshi S, Jothydev Kesavadev, Kumar P, Banshi Saboo, Mehta A, Bhattacharyya A, et al. Postprandial Glucose: A Variable in Continuum. Clinical Medicine Insights Endocrinology and Diabetes. 2025 Sep 1;18:11795514251370507-11795514251370507.
- Hershon KS, Hirsch BR, Odugbesan O. Importance of Postprandial Glucose in Relation to A1C and Cardiovascular Disease. Clinical Diabetes. 2019 Jul 1;37(3):250–9.
- Hulman A, Gujral UP, Narayan KMV, Pradeepa R, Mohan D, Anjana RM, et al. Glucose patterns during the OGTT and risk of future diabetes in an urban Indian population: The CARRS study. Diabetes Research and Clinical Practice. 2017 Apr;126:192–7.
- Lim MH, Chae H, Yoon J, Shin I. A deep learning framework for virtual continuous glucose monitoring and glucose prediction based on life-log data. Scientific Reports [Internet]. 2025 May 10;15(1). Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC12065870/
- Singh R, Marouane Toumi, Salathé M. Personalized glucose prediction using in situ data only. Frontiers in Nutrition. 2025 Jun 9;12.
- Ibáñez de Opakua A, Conde R, de Diego A, Bizkarguenaga M, Embade N, Lu SC, et al. Metabolomic-based aging clocks. npj Metabolic Health and Disease [Internet]. 2025 Sep 3 [cited 2026 Jan 12];3(1). Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC12408810/
- Li G, Cheng L, Wong IN, Yin Y, Chen J, Liu L, et al. Predicting healthspan and disease risks through biological age. Trends in Molecular Medicine [Internet]. 2025 Nov 28; Available from: https://www.sciencedirect.com/science/article/pii/S1471491425002576
- Wang K, Liu F, Wu W, Hu C, Shen X, Wang M, et al. A full life cycle biological clock based on routine clinical data and its impact in health and diseases. Nature Medicine [Internet]. 2025 Oct 27 [cited 2025 Oct 30]; Available from: https://www.nature.com/articles/s41591-025-04006-w?fromPaywallRec=false
- American Diabetes Association. Glycemic targets: Standards of medical care in diabetes—2022. Diabetes Care [Internet]. 2022 Dec 16;45(1):S83–96. Available from: https://diabetesjournals.org/care/article/45/Supplement_1/S83/138927/6-Glycemic-Targets-Standards-of-Medical-Care-in