Overeating as a 24-Hour Metabolic Stressor
Overeating in the evening can be conceptualized as a 24-hour metabolic stressor that perturbs both gastrointestinal physiology and circadian organization of glucose metabolism. Large, energy-dense meals slow gastric emptying and increase intragastric pressure, predisposing to gastroesophageal reflux when individuals subsequently lie supine. Nocturnal reflux events, in turn, are strongly associated with conscious awakenings and sleep fragmentation on polysomnography, particularly reductions in slow-wave sleep, which is normally protective against reflux by decreasing transient lower esophageal sphincter relaxation and suppressing arousal responses. Thus, even in the absence of overt gastroesophageal disease, eating close to bedtime can generate abdominal distension, heartburn, and bloating that disrupt continuity of sleep and reduce its restorative depth.
Beyond these mechanical consequences, high‑calorie, high‑carbohydrate evening meals impose a substantial metabolic load during a circadian phase in which glucose tolerance is intrinsically reduced and insulin secretion and sensitivity are declining. Laboratory protocols that compare identical test meals given in the biological morning versus evening consistently demonstrate higher postprandial glucose excursions at night, explained by attenuated early‑phase insulin release and relative insulin resistance, such that hyperglycemia and compensatory hyperinsulinemia may persist well into the habitual sleep period. This mismatch between sustained postprandial glycemic and insulinemic activity and the nocturnal down‑regulation of metabolic processes characterized by reduced energy expenditure and altered peripheral clock gene expression, positions late overeating as a driver of nocturnal glycemic volatility with downstream implications for arousals, autonomic activation, and next-day appetite regulation.
Late Eating, Circadian Misalignment, and Sleep Architecture
Late eating triggers profound circadian misalignment at the intersection of meal timing, sleep architecture, and glucose regulation, amplifying hunger, suppressing energy expenditure, and reprogramming adipose tissue metabolism toward fat storage even when total caloric intake remains constant. This cascade is strengthened by bidirectional interactions between sleep quality and glycemic control: late meals impair nocturnal sleep through mechanical and metabolic pathways, while fragmented or shortened sleep in turn worsens next-day glucose tolerance and intensifies carbohydrate cravings [1,2,3,4].
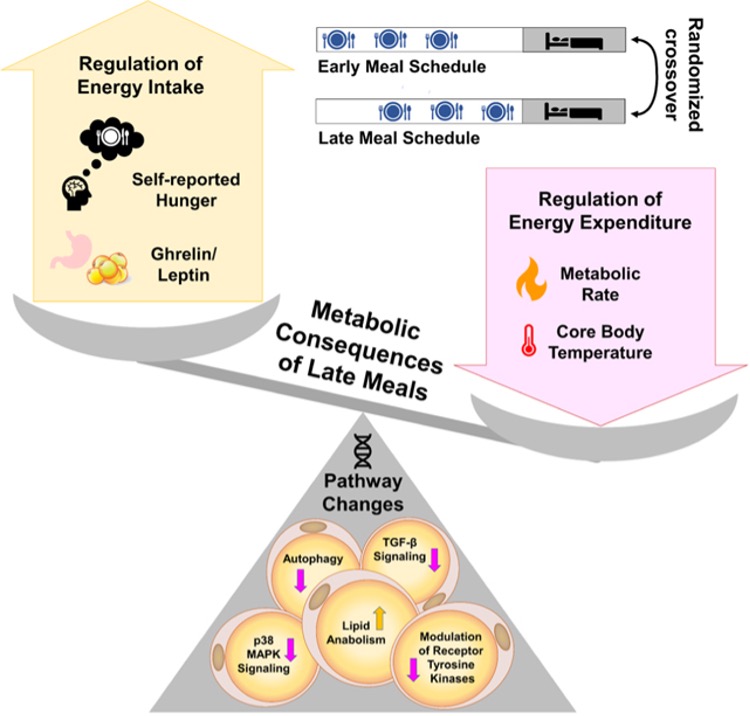
Randomized crossover trials in adults with overweight demonstrate that shifting identical meals approximately four hours later, from early eating (meals at 1, 5, and 9 hours after waking) to late eating (meals at 5, 9, and 13.5 hours after waking, with the final meal only 2.5 hours before sleep) produces striking effects on appetite regulation and energy balance that persist cross a full 24-hour cycle. In the landmark study by Vujović and colleagues, late eating doubled the probability of subjective hunger during waking hours (odds ratio 2.02, 95% confidence interval 1.46–3.35; p<0.0001) despite rigorous control of sleep duration, total caloric intake, macronutrient composition, physical activity, posture, and light exposure. This increased hunger was accompanied by a 6.3% reduction in waketime circulating leptin, the satiety hormone and a 34.24% rise in the waketime ghrelin:leptin ratio, indicating a persistent pro-hunger hormonal milieu. Across the full 24 hours, the ghrelin:leptin ratio remained elevated by 11.72% (p=0.006), reflecting a sustained shift toward orexigenic signalling that overlaps with the biological night and can destabilize sleep through nocturnal arousals and difficulty returning to sleep following awakenings [1,3,5].
Beyond appetite hormones, late eating decreased waketime energy expenditure by 4.94% (approximately 59 kilocalories per day; p=0.0002) and lowered 24-hour core body temperature by 0.19% (p=0.019), a validated proxy of total energy expenditure. Notably, core body temperature remained suppressed during the final four hours of the sleep episode, a period when acute diet-induced thermogenesis from the late evening meal would have dissipated, suggesting an enduring reduction in basal thermogenesis rather than a transient postprandial effect. This coordinated decrease in both measured energy expenditure and thermometry across wake and sleep periods indicates that late eating imposes a multi-hour metabolic penalty extending well into the following morning, effectively priming the organism for positive energy balance even before volitional food intake resumes [3,5].
At the molecular level, late eating induced coordinated changes in subcutaneous adipose tissue gene expression consistent with reduced lipolysis and increased adipogenesis. Pathway enrichment analyses of adipose biopsies collected at matched clock times in both early and late eating conditions revealed that late eating downregulated genes responsible for lipid breakdown, including phospholipase D6 (Ipld6) 2,4-dienoyl-CoA reductase (DERC1), acid ceramidase (ASAH1), and the lipolysis-promoting abhydrolase domain-containing protein 5 (ABHD5), while imultaneously upregulating lipogenic genes such as glycerol-3-phosphate acyltransferase (GPAM) ATP citrate lyase (ACLY) acetoacetyl-CoA synthetase (AACS) and ceramide kinase (CERK). Late eating also suppressed expression of genes in the p38 mitogen-activated protein kinase (MAPK) pathway, particularly growth arrest and DNA-damage-inducible gamma (GADD45g) a positive regulator of thermogenic energy expenditure in brown adipocytes; downregulated the transforming growth factor-β (TGF-β) signalling pathway, which normally inhibits adipogenesis; and reduced expression of autophagy-related genes, a profile that collectively favours adipocyte expansion and energy storage over mobilization [3,6,7].
These physiological and molecular changes occur without detectable differences in sleep duration or sleep stage distribution between early and late eating conditions, ruling out confounding by altered sleep quantity or architecture and confirming that meal timing exerts independent effects on metabolic regulation. The temporal alignment between elevated pro-hunger hormones during the biological night and the increased risk of nocturnal awakenings suggests a plausible mechanistic link: sustained elevation of the ghrelin:leptin ratio may not only drive next-day food intake but also fragment sleep continuity through direct effects on hypothalamic arousal circuits, thereby amplifying the metabolic cost of late eating through a secondary pathway of sleep disruption [1,8,9].
From Postprandial Peaks to Nocturnal Reactive Lows
High-glycemic, highly processed meals trigger a well-characterized sequence of rapid glucose elevation followed by exaggerated insulin secretion that can overshoot physiological requirements and drive blood glucose below baseline 2–5 hours later, a phenomenon termed postprandial reactive hypoglycemia. This pattern is particularly common in individuals with underlying insulin hyperresponsiveness, impaired first-phase insulin secretion, or insulin resistance, and is mechanistically linked to dysregulated incretin hormone signalling, delayed insulin kinetics, and insufficient glucagon counter regulation. The resulting hypoglycemic nadir is accompanied by a constellation of autonomic and neuroglycopenic symptoms that disrupt daily function and, critically, can fragment sleep if they occur during the first half of the night, thereby amplifying next-day metabolic dysfunction and carbohydrate-seeking behaviour [10,11,12,13,14,15,16].
Reactive hypoglycemia occurs when blood glucose falls to 55 mg/dL (approximately 3.0 mmol/L) or lower within 2–5 hours following food intake, and is clinically recognized in three temporally and mechanistically distinct subtypes: alimentary (early) hypoglycemia within 1–2 hours, idiopathic reactive hypoglycemia at approximately 3 hours, and late reactive hypoglycemia at 4–5 hours postprandially. The most common form, late reactive hypoglycemia arises from a loss of first-phase insulin secretion, an early metabolic defect also observed in impaired glucose tolerance and early type 2 diabetes. When first-phase insulin release is blunted or delayed, postprandial glucose rises more steeply and persists longer than normal, which subsequently triggers an exaggerated and inappropriately timed second-phase insulin response. This late but excessive insulin secretion continues even as intestinal glucose absorption wanes, creating a mismatch between circulating insulin levels and nutrient availability that drives blood glucose below baseline and sustains hyperinsulinemia for hours into the postabsorptive period [10,17,18,19].
High-glycemic-index carbohydrates and highly processed foods are particularly potent triggers of this insulin overshoot because they induce rapid and pronounced elevations in blood glucose often described colloquially as an “insulin spike” which the pancreas interprets as a demand for robust insulin secretion. In susceptible individuals, the body produces more insulin than is actually needed to handle the ingested carbohydrate load, and this surplus insulin persists after nutrients have been absorbed and cleared from the circulation, thereby lowering glucose concentrations below the fasting baseline. Sporadic consumption of high-carbohydrate snacks and meals throughout the day is recognized as the specific dietary pattern most likely to precipitate these “sugar crashes,” reinforcing a cycle in which rapid glycemic rises are followed by symptomatic lows [15,18,20,21].
The clinical presentation of reactive hypoglycemia reflects two overlapping physiological responses to declining glucose: autonomic (adrenergic) symptoms and neuroglycopenic symptoms. Autonomic symptoms including sweating, palpitations, tremor, shakiness, anxiety, pallor, tachycardia, and hunger, typically manifest when glucose falls below 70 mg/dL (approximately 3.9 mmol/L) and result from the body’s compensatory release of epinephrine and activation of the sympathetic nervous system. These symptoms serve as an early warning system, prompting individuals to seek food before more severe deficits occur. Neuroglycopenic symptoms such as fatigue, irritability, confusion, difficulty concentrating, drowsiness, weakness, incoordination, and impaired speech emerge when lucose drops below 45–50 mg/dL (2.5–2.8 mmol/L), reflecting direct deprivation of the brain’s primary fuel source. If untreated, neuroglycopenic symptoms can progress to severe confusion, loss of consciousness, and coma [22,23,24,25].
Importantly, reactive hypoglycemia during the nocturnal period poses unique risks for sleep disruption and prolonged metabolic compromise. Nocturnal hypoglycemia is associated with conscious awakenings, night sweats, damp clothing or sheets, headaches upon waking, nightmares, and impaired next-day mood and cognitive performance. Observational and experimental evidence demonstrates that even single episodes of nocturnal hypoglycemia adversely affect subjective well-being the following day, and recurrent exposure is implicated in the development of hypoglycemia unawareness, a dangerous syndrome in which autonomic warning symptoms are progressively blunted. In severe cases, nocturnal hypoglycemia has been linked to seizures, coma, and cardiac arrhythmias resulting in sudden death, the so-called “dead-in-bed syndrome” in diabetes [16,26,27].
Laboratory studies reveal that the awakening response to hypoglycemia during sleep is highly variable and frequently impaired. In a controlled trial by Schultes and colleagues, when blood glucose was lowered to 2.2 mmol/L (approximately 40 mg/dL) during stage 2 sleep, 62.5% of healthy control participants awakened, whereas only 6.25% of individuals with type 1 diabetes awakened at the same glycemic threshold. Critically, in all participants who did awaken, plasma epinephrine concentrations began to rise an average of 7.5 minutes before polysomnographic signs of wakefulness appeared, indicating that the awakening response is part of a centrally coordinated counterregulatory cascade launched in parallel with hormonal defense mechanisms. Failure to awaken during hypoglycemia increases the risk of prolonged, symptomatic, and potentially fatal episodes [14,28].
The interplay between sleep and hypoglycemia counterregulation is bidirectional: sleep itself raises the glycemic threshold at which counterregulatory hormones are released, meaning that lower glucose concentrations are tolerated during sleep than during wakefulness before epinephrine, cortisol, and glucagon secretion are triggered. Furthermore, sleep promotes adaptation to recurrent hypoglycemia, a process that, while potentially protective in the short term, can contribute to hypoglycemia unawareness over time. Conversely, sleep deprivation prevents this adaptation and preserves robust counterregulatory responses, suggesting that chronic sleep disruption may interact with glycemic volatility in complex, context-dependent ways [14,28].
The symptomatic experience of reactive hypoglycemia, particularly the combination of adrenergic arousal (palpitations, tremor, anxiety) and neuroglycopenic distress (fatigue, irritability, difficulty thinking) generates powerful cravings for fast-acting carbohydrates. Experimental evidence demonstrates that high-calorie, glucose-rich “comfort foods” rapidly attenuate neuroglycopenic symptoms, providing immediate subjective relief and thereby reinforcing carbohydrate-seeking behavior. This creates a vicious cycle: consumption of high-glycemic foods triggers insulin overshoot and reactive hypoglycemia, which in turn drives cravings for the same category of rapidly absorbed carbohydrates that initiated the glycemic swing. When these episodes occur during the first half of the night, the resulting sleep fragmentation amplifies next-day hunger, impairs glucose tolerance, and intensifies carbohydrate preference, perpetuating a 24-hour pattern of overeating, poor sleep, reactive lows, and intense cravings [1,3,20,29,30,31].
Reactive Hypoglycemia, Binge-Type Eating, and “Carb Echoes”
In cohorts with binge eating disorder and food addiction, the consumption of high-glycemic, hyper-palatable foods precipitates a specific metabolic and behavioral phenotype characterized by exaggerated reactive hypoglycemia and powerful carbohydrate-seeking behaviors that maintain disordered eating patterns through both biological and learned mechanisms. A large cross-sectional study of 200 adults with obesity and no diabetes demonstrated that individuals diagnosed with binge eating disorder or food addiction experienced reactive hypoglycemia, defined as symptomatic hypoglycemia (≤70 mg/dL) confirmed by Whipple’s triad (symptoms consistent with hypoglycemia, low plasma glucose, and symptom resolution with carbohydrate intake), at rates rates significantly exceeding those without eating pathology, and critically, the severity of food addiction was the strongest independent predictor of early and symptomatic hypoglycemic events during extended oral glucose tolerance testing [29].
In the study by Rania and colleagues, participants with binge eating disorder and food addiction (BED, FA, or comorbid BED-FA) underwent a 5-hour oral glucose tolerance test with continuous monitoring for hypoglycemic events and associated symptoms. Reactive hypoglycemia, which is hypoglycemia accompanied by neuroglycopenic or neurogenic symptoms such as tremor, sweating, palpitations, hunger, disorientation, impaired vision, or fainting was documented in 28% of the entire sample but showed striking variation by diagnostic group: 54.3% of participants with food addiction, 34.9% with binge eating disorder, 25.1% without eating pathology, and only 15.1% with comorbid BED-FA experienced reactive episodes (χ²=13.898, p=0.003). Importantly, the temporal profile of reactive hypoglycemia differed by diagnosis, with food addiction associated with earlier events (210–240 minutes post-glucose load) and binge eating disorder with later events (270 minutes), and time-to-event analysis confirmed that individuals with food addiction reached symptomatic hypoglycemia significantly faster than all other groups (log-rank χ²=13.952, p=0.003) [29].
Multivariate logistic regression adjusting for age, sex, body mass index, glucose tolerance profile, and insulin resistance (HOMA-IR) revealed that severity of food addiction as measured by the Yale Food Addiction Scale 2.0 was the only significant independent predictor of both early hypoglycemia occurring before 180 minutes) and reactive hypoglycemia, with individuals classified as having severe food addiction experiencing 14-fold higher odds of early hypoglycemia compared to those without food addiction (adjusted odds ratio 14.013, 95% CI 2.933–66.953; p=0.001). Notably, binge eating severity (assessed via the Binge Eating Scale) did not independently predict hypoglycemia outcomes after adjustment for confounders, although the BED group exhibited the highest fasting insulin and insulin resistance (HOMA-IR) values and was significantly more likely to experience severe hypoglycemia (glucose <54 mg/dL) at the late stage of the test (300 minutes; χ²=11.120, p=0.011) [29].
These findings support a bidirectional reinforcing loop in which repetitive consumption of high-glycemic-index, ultra-processed foods characteristic of binge eating and food addiction triggers exaggerated postprandial insulin secretion, particularly pronounced in individuals with insulin hyperresponsiveness, impaired first-phase insulin secretion, or heightened incretin effects leading to reactive hypoglycemia several hours later. The symptomatic hypoglycemic nadir, accompanied by autonomic arousal (palpitations, tremor, anxiety) and neuroglycopenic distress (fatigue, irritability, difficulty concentrating), generates intense hunger and specific cravings for the same category of rapidly absorbed carbohydrates that initiated the glycemic swing, thereby closing the loop and perpetuating the addictive eating pattern [10,20,29,32].
The biological substrate of this reinforcing loop involves the interaction between glucose, insulin, and mesolimbic dopamine reward circuitry. Mild hypoglycemia preferentially activates limbic-striatal brain regions including the nucleus accumbens, insula, and ventral tegmental area in response to visual food cues, producing greater subjective desire and “wanting” for high-calorie foods while simultaneously reducing activity in prefrontal executive control regions (prefrontal cortex, anterior cingulate cortex) that normally inhibit impulsive food consumption. Functional MRI studies using stepped hyperinsulinemic-hypoglycemic clamps demonstrate that transient modest reductions in circulating glucose increase striatal and insula activation to high-calorie food images by 20–30% and double the subjective wanting ratings for these foods, independent of changes in leptin or ghrelin but correlated with rises in cortisol—a stress hormone mobilized during hypoglycemia. This pattern reflects an adaptive survival mechanism in which hypoglycemia shifts neural control away from prefrontal inhibitory circuits and toward subcortical motivational and reward pathways, promoting food-seeking behaviour and preferentially increasing desire for energy-dense, glucose-rich foods that can rapidly restore circulating glucose [33,34,35,36].
Critically, insulin itself acts as a neuromodulator within striatal reward circuits, amplifying dopamine release through activation of insulin receptors on striatal cholinergic interneurons. Under normal conditions, acute insulin administration to the ventral tegmental area suppresses food intake and decreases motivation for palatable foods by enhancing dopamine signaling and autoreceptor-mediated inhibition of dopamine neuron firing. However, chronic consumption of high-glycemic foods and repeated cycles of hyperinsulinemia and reactive hypoglycemia lead to insulin resistance within these same reward circuits, paradoxically increasing limbic-striatal activation in response to food cues and promoting compulsive eating. Obese individuals and those with insulin resistance show greater activation of limbic regions (amygdala, striatum, orbitofrontal cortex, insula) when exposed to food stimuli compared to insulin-sensitive controls, and exhibit reduced availability of striatal D2 dopamine receptors, a profile consistently associated with compulsive food intake and diminished metabolic activity in prefrontal executive control regions [37,38,39,40,41,42].
This neurobiological cascade creates a powerful behavioural echo: hypoglycemia triggers craving for high-glycemic foods through direct action on reward and arousal circuits; consumption of these foods provides immediate symptomatic relief and dopamine-mediated reward reinforcement; the resulting glucose-insulin overshoot precipitates a subsequent hypoglycemic episode; and the cycle repeats with increasing intensity as insulin resistance develops and dopaminergic reward sensitivity blunts, requiring progressively larger food intake to achieve equivalent hedonic response. As Rania and colleagues propose, “reactive hypoglycemia, supported by glucose and insulin fluctuations, may reinforce food intake and disordered eating behaviours (e.g., binge eating, food addiction) which in turn may exacerbate reactive hypoglycemia.” This self-perpetuating mechanism operates both biologically, through metabolic amplification of glucose-insulin swings and dopaminergic reward sensitization and behaviourally through learned associations between hypoglycemic distress, carbohydrate consumption, and symptomatic relief that condition future carbohydrate-seeking responses [29,32,33,37,39].
When these reactive hypoglycemic episodes occur during the first half of the night following an evening of overeating, the resulting autonomic and neuroglycopenic symptoms fragment sleep continuity, reduce slow-wave sleep, and amplify next-day hunger, insulin resistance, and carbohydrate preference, thereby extending the metabolic and behavioural dysfunction across the full 24-hour cycle and manifesting as powerful morning and daytime carb-seeking behaviour, the “carb echo” of nocturnal glycemic instability [1,3,29,30,31,33].
Sleep Loss, Appetite Hormones, and Next-Day Intake
Experimental sleep restriction studies conducted under rigorous laboratory conditions consistently demonstrate that curtailed or fragmented sleep profoundly disrupts appetite-regulating hormones, shifting the endocrine milieu toward increased hunger, heightened preference for energy-dense and carbohydrate-rich foods, and substantial elevations in total caloric intake, even in the absence of changes in energy expenditure. These findings provide mechanistic insight into the well-documented epidemiological association between short sleep duration and obesity risk, and establish sleep as a powerful, modifiable driver of energy balance and macronutrient selection [43,44,45,46,47,48,49].
Meta-analytic evidence pooling results from 11 randomized controlled and crossover trials with 172 participants reveals that partial sleep deprivation, defined as sleeping fewer than 6.5 hours per night, significantly increases energy intake by 385 kilocalories per day (95% confidence interval 252–517 kcal; p<0.00001) compared to normal sleep conditions, with no compensatory increase in total energy expenditure or resting metabolic rate, thereby creating a positive energy balance of approximately 385 kcal/day that, if sustained, would be sufficient to drive meaningful weight gain over time. Additional meta-analyses confirm these findings, reporting energy intake increases ranging from 150 to 385 kcal per day with sleep restriction, accompanied by higher consumption of dietary fat and reduced protein intake, although effects on total carbohydrate intake are inconsistent across studies when expressed as a proportion of total energy [45,46,48,50,51].
The hormonal substrate underlying this increased appetite involves complex and sometimes paradoxical changes in circulating leptin and ghrelin. The landmark study by Spiegel and colleagues demonstrated that restricting sleep to 4 hours per night for 6 nights in healthy young men reduced mean circulating leptin by 19%, maximal leptin by 26%, and leptin rhythm amplitude by 20% compared to extended sleep of 12 hours per night under controlled feeding conditions, while simultaneously increasing ghrelin by 28%. These hormonal shifts accounted for approximately 70% of the variance in increased hunger ratings during sleep restriction, with participants reporting 24% higher hunger and 23% greater appetite during short sleep, and showing particularly elevated cravings for carbohydrate-rich foods such as sweets, salty snacks, and starchy foods [44,52,53].
However, the direction and magnitude of leptin and ghrelin responses to sleep restriction vary considerably depending on energy balance status, sex, body mass index, and whether participants have ad libitum access to food or receive controlled meals. Studies that allow free food access during sleep restriction paradoxically report increased leptin levels, particularly morning leptin, while those that maintain strict caloric control or negative energy However, the direction and magnitude of leptin and ghrelin responses to sleep restriction vary considerably depending on energy balance status, sex, body mass index, and whether participants have ad libitum access to food or receive controlled meals. Studies that allow free food access during sleep restriction paradoxically report increased leptin levels [44,52,53].
Ghrelin responses to sleep restriction show more consistency, with most studies reporting elevated circulating ghrelin concentrations during both nocturnal and daytime periods following sleep loss. In a frequently sampled 24-hour hormonal profiling study, Broussard and colleagues found that restricting sleep to 4.5 hours for 4 nights significantly increased total ghrelin area-under-the-curve by 10% compared to 8.5 hours of sleep (p<0.01), with postprandial ghrelin levels remaining elevated even after breakfast, lunch, and dinner meals. When participants were subsequently given ad libitum access to food, sleep restriction increased total caloric intake from snacks by 328±140 kcal (p=0.03), driven primarily by a rise in carbohydrate consumption (p=0.02) rather than fat or protein, and critically, the magnitude of evening ghrelin elevation during sleep restriction correlated significantly with greater consumption of calories from sweets (r=0.48, p=0.04), establishing ghrelin as a predictive biomarker of carbohydrate-seeking behaviour following sleep loss [44,49].
The specificity of sleep restriction’s effects on carbohydrate intake particularly snacks, desserts, and sweets is further corroborated by pediatric and adolescent studies. In a crossover trial of adolescents, experimentally restricting sleep to approximately 6.5 hours for four consecutive nights compared to 9 hours of sleep resulted in significantly higher dietary glycemic index and glycemic load the following day, with a trend toward greater total caloric and carbohydrate intake and a marked increase in consumption of desserts and sweets, while fat and protein intakes remained unchanged. Similarly, cross-sectional data in children aged 9–11 years demonstrate that shorter sleep duration is associated with higher carbohydrate intake, and experimental sleep manipulation studies consistently show that sleep-deprived children consume 42–134 fewer calories per day when sleep is extended by 1.5–3 hours, with the reduction driven primarily by decreased carbohydrate-rich snack consumption [47,50,54].
Recent meta-analyses further refine our understanding of the temporal and compositional patterns of increased intake following sleep restriction. González-Ortiz and colleagues found that partial sleep deprivation increases not only total energy intake by approximately 150 kcal per day (95% CI 10–290; p=0.04) but also consumption of all three macronutrients which includes carbohydrates, fats, and protein when analysed in absolute rather than relative terms. Importantly, the increase in food intake is not evenly distributed across the day: multiple studies document that sleep-restricted individuals consume significantly more calories from snacks particularly evening and nighttime snacks compared to daytime meals, with one study reporting that snack consumption increased by 30% during sleep restriction, while breakfast, lunch, and dinner intake remained relatively stable [44,51,54,55].
The interaction between sleep restriction and carbohydrate quality further modulates these effects. Cross-sectional analyses using continuous glucose monitoring demonstrate that individuals consuming higher quantities of low-quality carbohydrates (high glycemic index, refined grains, added sugars) have 39% higher odds of experiencing poor sleep patterns (OR 1.39, 95% CI 1.20–1.61), while those consuming more high-quality carbohydrates (whole grains, fruits, non-starchy vegetables) have 29% lower odds of poor sleep (OR 0.71, 95% CI 0.62–0.81), suggesting a bidirectional relationship in which poor sleep increases craving for low-quality carbohydrates, and consumption of these foods in turn worsens subsequent sleep quality, creating a self-perpetuating cycle [44,56].
Notably, the hormonal and behavioural effects of sleep restriction can be partially reversed with sleep extension. A large randomized clinical trial of 80 adults with overweight and habitual sleep duration <6.5 hours per night demonstrated that a 2-week sleep extension intervention which increased average sleep duration by approximately 1.2 hours per night, significantly reduced daily energy intake by 270 kcal compared to controls (who increased intake by 115 kcal; p<0.001), with each additional hour of sleep associated with a 162 kcal reduction in energy intake (p<0.001), and resulting in a clinically meaningful negative energy balance without any dietary counselling or caloric restriction instructions. These findings establish that sleep duration exerts a bidirectional, dose-dependent control over energy intake: restriction increases consumption, while extension decreases it, independent of volitional dietary changes [44,57].
Taken together, the experimental literature demonstrates that sleep restriction of even modest duration (4–5 hours for 4–6 nights) consistently increases total caloric intake by 250–550 kcal per day in tightly controlled laboratory settings, with the excess calories derived disproportionately from carbohydrate-rich snacks, particularly sweets and desserts consumed during evening hours and predicted by elevations in ghrelin and, under ad libitum conditions, paradoxical rises in leptin reflecting positive energy balance. This robust pattern, replicated across diverse populations including healthy lean men, women, children, adolescents, and adults with overweight, underscores sleep as a fundamental regulator of appetite, macronutrient preference, and energy balance, with direct relevance to the overeating-sleep-craving cycle targeted by metabolic wellness interventions [44,45,47,49].
Neurocognitive Bias Toward High-Reward Carbohydrates
Sleep restriction creates a neurocognitive bias toward high-reward, high-carbohydrate foods by amplifying brain reward responses while weakening executive control over eating. After several nights of curtailed sleep, fMRI studies show stronger activation to images of high-sugar, high-fat foods in limbic-striatal and orbitofrontal regions, alongside reduced engagement of prefrontal areas that normally help inhibit impulsive choices [44,59,60,61].
Concurrently, sleep loss heightens circulating endocannabinoids such as 2‑arachidonoylglycerol, whose daily rhythm becomes amplified and shifted later into the day, paralleling increased hunger and snack desire. In ad libitum settings, this neurochemical and neural profile leads to greater intake of calorie‑dense snacks, especially sweets and refined carbohydrates despite standardized meals, linking inadequate sleep directly to next day preference for quick energy, high glycemic foods [44,49,62,63].
Carbohydrate Intake, Sleep Quality, and a Bidirectional Loop
Carbohydrate intake and sleep quality appear to form a bidirectional loop in which diet influences sleep and sleep, in turn, shapes carbohydrate choices. Cross-sectional and cohort data indicate that higher total carbohydrate load, especially from low quality, high glycemic sources is associated with greater odds of insomnia, poorer subjective sleep quality and more irregular sleep patterns, whereas moderate intake of high0quality carbohydrates (higher fiber, lower glycemic load) correlates with better sleep and mental health outcomes, particularly in older adults [44,47,56,64,65].
Experimental work then shows the opposite direction of effect: even a few nights of delayed bedtime or restricted sleep in adolescents and young adults reliably increase dietary glycemic index and glycemic load the next day, with a specific rise in desserts, sweets, and sugar‑sweetened beverages and little change in fat or protein intake. Narrative reviews on sleep and eating behaviour synthesize these findings into a feedback model where low‑quality, high‑glycemic diets fragment sleep via glycemic volatility and nocturnal arousals, and poor or curtailed sleep subsequently amplifies hedonic drive for high‑sugar, high‑GI foods, thereby perpetuating a 24‑hour cycle of disturbed sleep and carbohydrate‑seeking behaviour [44,47,66,67,68].
Practical Implications
Evening overeating emerges as a clinically relevant target for 24‑hour metabolic regulation, with implications for both sleep quality and appetite control the following day. The converging data suggest that adjusting when, what, and how much is eaten in the latter part of the day can meaningfully reduce nocturnal glycaemic volatility and dampen next‑day carbohydrate‑seeking behaviour in at‑risk populations
Practical Implications
From a clinical and programmatic perspective, large, energy‑dense evening meals rich in refined starches and added sugars should be conceptualized as a modifiable driver of reactive hypoglycaemia and compensatory overeating rather than a benign habit. High‑glycaemic loads late in the day promote large postprandial glucose and insulin excursions, followed by sub‑ or overt hypoglycaemic dips that are associated with autonomic symptoms, increased hunger, and higher frequency of snacking and binge‑type intake.
Pragmatically, metabolic wellness and healthy aging programs can incorporate three simple levers: moderating evening portion size, upgrading carbohydrate quality, and shifting energy intake earlier. Emphasizing lower‑GI, higher‑fibre carbohydrate sources paired with protein and healthy fats in the last meal can blunt postprandial glycaemic peaks and reduce the risk of late reactive hypoglycaemia, while also improving satiety. In parallel, operationalizing an “evening energy cut‑off” (for example, limiting caloric intake in the 3–4 hours before habitual bedtime) aligns feeding with circadian patterns of insulin sensitivity and may reduce nocturnal reflux and arousals, indirectly supporting more consolidated sleep.
Early time‑restricted eating (eTRE), in which caloric intake is compressed into an earlier 6–10‑hour window, represents a feasible structure for many adults and has been shown to improve 24‑hour glucose profiles, insulin sensitivity, body weight, and blood pressure when combined with caloric restriction. For patients already engaged in weight‑loss or cardiometabolic programs, layering an earlier eating window onto existing dietary prescriptions may therefore offer an additional, low‑cost means of stabilizing nocturnal glucose and attenuating the “carb crash–craving” cycle without adding pharmacological complexity.
Conclusion
Taken together, current evidence supports a mechanistic sequence in which evening overeating, particularly of refined carbohydrates and fats, induces large postprandial glucose and insulin swings, predisposes to nocturnal or early morning reactive hypoglycemia, nd perturbs sleep via both mechanical and autonomic pathways. These sleep disruptions, in turn, are consistently linked to elevations in ghrelin, reductions in leptin, and increased intake of energy‑dense, carbohydrate‑rich snacks, providing a plausible endocrine and behavioural bridge to next‑day cravings for fast‑acting carbohydrates.
Within metabolic wellness and healthy aging frameworks, moderating evening portion sizes, improving the quality and glycaemic profile of evening carbohydrates, and preferentially advancing caloric intake toward the earlier part of the day through strategies such as eTRE constitute evidence-aligned, scalable interventions to stabilize nocturnal glucose, protect sleep architecture, and break the recurrent “carb crash–craving” loop across the 24‑hour period. Framing these adjustments as core components of circadian‑aligned nutrition, rather than restrictive “rules” about late‑night eating, may enhance adherence and facilitate integration into digital, AI‑supported metabolic health programs.
Reference
- Chaput JP, McHill AW, Cox RC, Broussard JL, Dutil C, da Costa BGG, et al. The role of insufficient sleep and circadian misalignment in obesity. Nature Reviews Endocrinology. 2022 Oct 24;19(2).
- Mohd D. Association Between Sleep Quality and Glycemic Variability in Patients with Type 2 Diabetes Mellitus: A Prospective Study. European Journal of Cardiovascular Medicine [Internet]. 2026 Jan 2 [cited 2026 Jan 6];16:30–5. Available from: https://www.healthcare-bulletin.co.uk/article/association-between-sleep-quality-and-glycemic-variability-in-patients-with-type-2-diabetes-mellitus-a-prospective-study-4718/
- Vujović N, Piron MJ, Qian J, Chellappa SL, Nedeltcheva A, Barr D, et al. Late isocaloric eating increases hunger, decreases energy expenditure, and modifies metabolic pathways in adults with overweight and obesity. Cell Metabolism [Internet]. 2022 Oct 4;34(10):1486-1498.e7. Available from: https://www.cell.com/cell-metabolism/fulltext/S1550-4131(22)00397-7?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1550413122003977%3Fshowall%3Dtrue
- Jung H, Choung RS, Talley NJ. Gastroesophageal Reflux Disease and Sleep Disorders: Evidence for a Causal Link and Therapeutic Implications. Journal of Neurogastroenterology and Motility [Internet]. 2010 Jan 31;16(1):22–9. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2879818/
- Harris M. RCT Results: Eating Later in the Day and Impact on Hunger and Energy Expenditure – The ObG Project [Internet]. The ObG Project. ObG Project; 2022 [cited 2026 Jan 6]. Available from: https://www.obgproject.com/2022/10/23/rct-results-eating-later-in-the-day-and-impact-on-hunger-and-energy-expenditure/
- Peters B, Vahlhaus J, Pivovarova-Ramich O. Meal timing and its role in obesity and associated diseases. Frontiers in Endocrinology. 2024 Mar 22;15.
- BaHammam AS, Pirzada R. Timing matters: The interplay between early mealtime, circadian rhythms, gene expression, circadian hormones, and metabolism—a narrative review. Clocks & sleep [Internet]. 2023 Sep 6;5(3):507–35. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC10528427/
- Kim TW, Jeong JH, Hong SC. The Impact of Sleep and Circadian Disturbance on Hormones and Metabolism. International Journal of Endocrinology [Internet]. 2015 Mar 11;2015(591729):1–9. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4377487/
- Bodosi B, Gardi J, Hajdu I, Szentirmai E, Obal F, Krueger JM. Rhythms of ghrelin, leptin, and sleep in rats: effects of the normal diurnal cycle, restricted feeding, and sleep deprivation. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2004 Nov;287(5):R1071–9.
- Altuntaş Y. Postprandial Reactive Hypoglycemia. Şişli Etfal Hastanesi tıp Bülteni [Internet]. 2019 Aug 28;53(3):215–20. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7192270/
- Castro R. Reactive hypoglycemia: What causes it? [Internet]. Mayo Clinic. 2016. Available from: https://www.mayoclinic.org/diseases-conditions/diabetes/expert-answers/reactive-hypoglycemia/faq-20057778
- Reactive hypoglycemia: Causes, symptoms, and treatment [Internet]. www.medicalnewstoday.com. 2021. Available from: https://www.medicalnewstoday.com/articles/reactive-hypoglycemia
- How can a person avoid nocturnal hypoglycemia? [Internet]. www.medicalnewstoday.com. 2023. Available from: https://www.medicalnewstoday.com/articles/nocturnal-hypoglycemia
- Schultes B, Jauch-Chara K, Gais S, Hallschmid M, Reiprich E, Kern W, et al. Defective Awakening Response to Nocturnal Hypoglycemia in Patients with Type 1 Diabetes Mellitus. Groop LC, editor. PLoS Medicine. 2007 Feb 27;4(2):e69.
- Brun JF, Fedou C, Mercier J. Postprandial reactive hypoglycemia. Diabetes & Metabolism [Internet]. 2000 Nov 1;26(5):337–51. Available from: https://pubmed.ncbi.nlm.nih.gov/11119013/
- Allen KV, Frier BM. Nocturnal Hypoglycemia: Clinical Manifestations and Therapeutic Strategies Toward Prevention. Endocrine Practice. 2003 Nov;9(6):530–43.
- Ahmed FW, Majeed MS, Kirresh O. Non-diabetic Hypoglycemia [Internet]. PubMed. Treasure Island (FL): StatPearls Publishing; 2023. Available from: https://www.ncbi.nlm.nih.gov/books/NBK573079/
- Kandel D, Bojsen-Møller KN, Svane MS, Samkani A, Astrup A, Holst JJ, et al. Mechanisms of action of a carbohydrate-reduced, high-protein diet in reducing the risk of postprandial hypoglycemia after Roux-en-Y gastric bypass surgery. The American journal of clinical nutrition [Internet]. 2019 Jan;110(2):296–304. Available from: https://pubmed.ncbi.nlm.nih.gov/30624666/
- Delacruz L, Adamczak LJ, Astapova O. Postprandial Hypoglycemia Associated With Insulin ReceptorGene Variant G1146R. JCEM Case Reports [Internet]. 2025 Sep 25 [cited 2026 Jan 6];3(12):luaf247–7. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC12559848/
- Blood Sugar Metabolism Disorders: Hypoglycemia [Internet]. Restorative Medicine. 2025 [cited 2026 Jan 6]. Available from: https://restorativemedicine.org/fundamentals-naturopathic-endocrinology/blood-sugar-metabolism-disorders/hypoglycemia/
- Prakash K, Kabadi M, Kabadi UM. Idiopathic Reactive Hypoglycemia: Mechanisms of Onset and Remission with High Protein Low Carbohydrate Diet. Open Journal of Endocrine and Metabolic Diseases. 2015;05(09):117–23.
- LinkedIn [Internet]. Linkedin.com. 2026 [cited 2026 Jan 6]. Available from: https://www.linkedin.com/pulse/hypoglycemia-symptoms-treatment-baljinder-singh-1hiqc
- Rosenfeld E, Thornton PS. Hypoglycemia in Neonates, Infants, and Children [Internet]. PubMed. South Dartmouth (MA); 2023. Available from: https://www.ncbi.nlm.nih.gov/books/NBK594592/
- McMaster Textbook of Internal Medicine [Internet]. Empendium.com. 2024. Available from: https://empendium.com/mcmtextbook-sae/chapter/B78.II.24.10.?rfmcm
- Xu C, Yogaratnam J, Lua R, Naik S, Khoo CL, Pillai SS, et al. Persistent, severe hypoglycemia-induced organic brain syndrome with neurological sequelae: a case report. General Hospital Psychiatry [Internet]. 2011 Jun 11;33(4):412.e9–11. Available from: https://www.sciencedirect.com/science/article/abs/pii/S0163834311001794
- How can a person avoid nocturnal hypoglycemia? [Internet]. www.medicalnewstoday.com. 2023. Available from: https://www.medicalnewstoday.com/articles/nocturnal-hypoglycemia
- Hypoglycemia: Nocturnal | Johns Hopkins Diabetes Guide [Internet]. Hopkinsguides.com. 2016 [cited 2026 Jan 6]. Available from: https://www.hopkinsguides.com/hopkins/view/Johns_Hopkins_Diabetes_Guide/547068/7/Hypoglycemia:_Nocturnal
- Meyhöfer S, Dembinski K, Schultes B, Born J, Wilms B, Lehnert H, et al. Sleep deprivation prevents counterregulatory adaptation to recurrent hypoglycaemia. Diabetologia. 2022 Apr 21;
- Rania M, Caroleo M, Elvira Anna Carbone, Ricchio M, Maria Chiara Pelle, Zaffina I, et al. Reactive hypoglycemia in binge eating disorder, food addiction, and the comorbid phenotype: unravelling the metabolic drive to disordered eating behaviours. Journal of eating disorders. 2023 Sep 19;11(1).
- Maloney A, Kanaley JA. Short Sleep Duration Disrupts Glucose Metabolism: Can Exercise Turn Back the Clock? Exercise and Sport Sciences Reviews. 2024 Apr 12;52(3):77–86.
- Mohd D. Association Between Sleep Quality and Glycemic Variability in Patients with Type 2 Diabetes Mellitus: A Prospective Study. European Journal of Cardiovascular Medicine [Internet]. 2026 Jan 2;16:30–5. Available from: https://www.healthcare-bulletin.co.uk/article/association-between-sleep-quality-and-glycemic-variability-in-patients-with-type-2-diabetes-mellitus-a-prospective-study-4718/
- Klement J, Hubold C, Cords H, Oltmanns KM, Hallschmid M, Born J, et al. High-Calorie Glucose-Rich Food Attenuates Neuroglycopenic Symptoms in Patients with Addison’s Disease. The Journal of Clinical Endocrinology & Metabolism. 2010 Feb;95(2):522–8.
- Page KA, Seo D, Belfort-DeAguiar R, Lacadie C, Dzuira J, Naik S, et al. Circulating glucose levels modulate neural control of desire for high-calorie foods in humans. Journal of Clinical Investigation. 2011 Oct 3;121(10):4161–9.
- Frank S, Veit R, Sauer H, Enck P, Friederich HC, Unholzer T, et al. Dopamine Depletion Reduces Food-Related Reward Activity Independent of BMI. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology [Internet]. 2016 May 1;41(6):1551–9. Available from: https://pubmed.ncbi.nlm.nih.gov/26450814/
- Aklan I, Sayar Atasoy N, Yavuz Y, Ates T, Coban I, Koksalar F, et al. NTS Catecholamine Neurons Mediate Hypoglycemic Hunger via Medial Hypothalamic Feeding Pathways. Cell Metabolism [Internet]. 2020 Feb 4 [cited 2021 Nov 3];31(2):313-326.e5. Available from: https://www.sciencedirect.com/science/article/pii/S1550413119306230
- Blum K, Thanos PK, Gold MS. Dopamine and glucose, obesity, and reward deficiency syndrome. Frontiers in Psychology. 2014 Sep 17;5.
- Volkow ND, Wang GJ, Baler RD. Reward, Dopamine and the Control of Food intake: Implications for Obesity. Trends in Cognitive Sciences [Internet]. 2011 Jan;15(1):37–46. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3124340/
- Gruber J, Hanssen R, Mishal Qubad, Aicha Bouzouina, Schack V, Sochor H, et al. Impact of insulin and insulin resistance on brain dopamine signalling and reward processing – An underexplored mechanism in the pathophysiology of depression? Neuroscience & biobehavioral reviews/Neuroscience and biobehavioral reviews. 2023 Jun 1;149:105179–9.
- Arora K. How does dopamine impact blood glucose? [Internet]. Ultrahuman. 2022 [cited 2026 Jan 6]. Available from: https://blog.ultrahuman.com/blog/how-dopamine-impacts-blood-glucose/
- Frank S, Veit R, Sauer H, Enck P, Friederich HC, Unholzer T, et al. Dopamine Depletion Reduces Food-Related Reward Activity Independent of BMI. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology [Internet]. 2016 May 1;41(6):1551–9. Available from: https://pubmed.ncbi.nlm.nih.gov/26450814/
- Stouffer MA, Woods CA, Patel JC, Lee CR, Witkovsky P, Bao L, et al. Insulin enhances striatal dopamine release by activating cholinergic interneurons and thereby signals reward. Nature Communications. 2015 Oct 27;6(1).
- Interplay of Dopamine and Food Signals: Examining the Interaction of Dopamine and Homeostatic Signals in Food-motivated Behavior | Auctores [Internet]. Auctores. 2024 [cited 2026 Jan 6]. Available from: https://www.auctoresonline.org/article/interplay-of-dopamine-and-food-signals-examining-the-interaction-of-dopamine-and-homeostatic-signals-in-food-motivated-behavior
- Ad C, Re C, T A, Pg M, Fn A, C van der W, et al. Effects of Experimental Sleep Restriction on Caloric Intake and Activity Energy Expenditure [Internet]. Chest. 2013. Available from: https://pubmed.ncbi.nlm.nih.gov/23392199/
- St-Onge MP. The Role of Sleep Duration in the Regulation of Energy Balance: Effects on Energy Intakes and Expenditure. Journal of Clinical Sleep Medicine. 2013 Jan 15;
- Al Khatib HK, Harding SV, Darzi J, Pot GK. The effects of partial sleep deprivation on energy balance: a systematic review and meta-analysis. European Journal of Clinical Nutrition. 2016 Nov 2;71(5):614–24.
- Sleep deprivation may cause people to eat more calories | Website archive | King’s College London [Internet]. Kcl.ac.uk. 2017. Available from: https://www.kcl.ac.uk/archive/news/kings/newsrecords/2016/11 november-/sleep-deprivation-may-cause-people-to-eat-more-calories
- Beebe DW, Simon S, Summer S, Hemmer S, Strotman D, Dolan LM. Dietary Intake Following Experimentally Restricted Sleep in Adolescents. Sleep. 2013 Jun 1;36(6):827–34.
- Zhu B, Shi C, Park CG, Zhao X, Reutrakul S. Effects of sleep restriction on metabolism-related parameters in healthy adults: A comprehensive review and meta-analysis of randomized controlled trials. Sleep Medicine Reviews. 2019 Jun;45:18–30.
- Broussard JL, Kilkus JM, Delebecque F, Abraham V, Day A, Whitmore HR, et al. Elevated ghrelin predicts food intake during experimental sleep restriction. Obesity. 2015 Oct 15;24(1):132–8.
- Soltanieh S, Solgi S, Ansari M, Santos HO, Abbasi B. Effect of sleep duration on dietary intake, desire to eat, measures of food intake and metabolic hormones: A systematic review of clinical trials. Clinical Nutrition ESPEN. 2021 Oct;45:55–65.
- Espinosa Cuevas Á, González-Ortiz A, López-Bautista F, Valencia-Flores M. Effects of partial sleep deprivation on dietary energy intake in a healthy population: a systematic review and meta-analysis. Nutrición Hospitalaria. 2020;
- Knutson KL. Impact of Sleep and Sleep Loss on Glucose Homeostasis and Appetite Regulation. Sleep Medicine Clinics. 2007 Jun;2(2):187–97.
- Taheri S, Lin L, Austin D, Young T, Mignot E. Short Sleep Duration Is Associated with Reduced Leptin, Elevated Ghrelin, and Increased Body Mass Index. Froguel P, editor. PLoS Medicine [Internet]. 2004 Dec 7;1(3):e62. Available from: https://journals.plos.org/plosmedicine/article?id=10.1371/journal.pmed.0010062
- Martinez SM, Tschann JM, Butte NF, Gregorich SE, Penilla C, Flores E, et al. Short Sleep Duration Is Associated With Eating More Carbohydrates and Less Dietary Fat in Mexican American Children. Sleep [Internet]. 2016 Dec 9 [cited 2019 Oct 21];40(2). Available from: https://academic.oup.com/sleep/article/40/2/zsw057/2662318
- Tajiri E, Yoshimura E, Tobina T, Yamashita T, Kume K, Hatamoto Y, et al. Effects of sleep restriction on food intake and appetite under free-living conditions: A randomized crossover trial. Appetite [Internet]. 2023 Oct 1 [cited 2024 May 10];189:106998. Available from: https://pubmed.ncbi.nlm.nih.gov/37562755/
- Zhao Y, Guo H. The relationship between carbohydrate intake and sleep patterns. Frontiers in Nutrition. 2024 Dec 4;11.
- Tasali E, Wroblewski K, Kahn E, Kilkus J, Schoeller DA. Effect of Sleep Extension on Objectively Assessed Energy Intake Among Adults With Overweight in Real-life Settings. JAMA Internal Medicine. 2022 Feb 7;182(4).
- Ad C, Re C, T A, Pg M, Fn A, C van der W, et al. Effects of Experimental Sleep Restriction on Caloric Intake and Activity Energy Expenditure [Internet]. Chest. 2013. Available from: https://pubmed.ncbi.nlm.nih.gov/23392199/
- Demos KE, Sweet LH, Hart CN, McCaffery JM, Williams SE, Mailloux KA, et al. The Effects of Experimental Manipulation of Sleep Duration on Neural Response to Food Cues. Sleep. 2017 Jul 20;40(11).
- Satterfield BC, Raikes AC, Killgore WDS. Rested-Baseline Responsivity of the Ventral Striatum Is Associated With Caloric and Macronutrient Intake During One Night of Sleep Deprivation. Frontiers in Psychiatry. 2019 Jan 17;9.
- Kingshott R. MRI scans show how sleep loss affects the ability to choose proper foods – American Academy of Sleep Medicine – Association for Sleep Clinicians and Researchers [Internet]. American Academy of Sleep Medicine – Association for Sleep Clinicians and Researchers. 2012 [cited 2026 Jan 6]. Available from: https://aasm.org/mri-scans-show-how-sleep-loss-affects-the-ability-to-choose-proper-foods/
- Sleep loss boosts hunger and high-calorie food choices | University of Chicago News [Internet]. news.uchicago.edu. 2016. Available from: https://news.uchicago.edu/story/sleep-loss-boosts-hunger-and-high-calorie-food-choices
- Hanlon EC, Tasali E, Leproult R, Stuhr KL, Doncheck E, de Wit H, et al. Sleep Restriction Enhances the Daily Rhythm of Circulating Levels of Endocannabinoid 2-Arachidonoylglycerol. Sleep. 2016 Mar 1;39(3):653–64.
- Ghosn B, Abbasi H, Dehnavi MK, Hajian PN, Azadbakht L. The association between quality and quantity of carbohydrate with sleep, mood, anxiety, depression and stress among elderly. BMC Geriatrics. 2025 Jun 2;25(1).
- Farhadnejad H, Sadat S, Jahromi MK, Teymoori F, Tehrani AN, Mokhtari E, et al. The association of dietary glycemic index and glycemic load with the risk of insomnia in the adult population. BMC Nutrition. 2023 Feb 7;9(1).
- Duraccio KM, Whitacre C, Krietsch KN, Zhang N, Summer S, Price M, et al. Losing sleep by staying up late leads adolescents to consume more carbohydrates and a higher glycemic load. Sleep. 2021 Dec 17;45(3).
- Cheng WY, Chan WS. A Narrative Review on Sleep and Eating Behavior. Current Diabetes Reports. 2025 Sep 30;25(1).
- Simon SL, Field J, Miller LE, DiFrancesco M, Beebe DW. Sweet/Dessert Foods Are More Appealing to Adolescents after Sleep Restriction. Mistlberger RE, editor. PLOS ONE. 2015 Feb 23;10(2):e0115434.